Directeur de Recherche émérite au CNRS, Yves Carton a travaillé comme généticien au Laboratoire Evolution, Génomes, Comportement, Ecologie du CNRS et de l’Université Paris-Saclay (EGCE). Depuis plusieurs années, il se consacre à l’histoire des sciences, en particulier dans le domaine de l’entomologie et du darwinisme.
Passion-Entomologie souhaite vivement remercier M. Carton d’avoir accepté de répondre à ces quelques questions.
Vous êtes Directeur de Recherche émérite au CNRS, diplômé en immunologie et en sérologie, et vous avez travaillé sur l’insecte. Pouvez-vous nous expliquer en quoi consistait votre travail et quelles étaient les thématiques abordées ?
Yves Carton – Directeur de Recherche émérite au CNRS (Source : Y. Carton)
J’ai été très tôt, au cours de ma thèse, intéressé par l’étude des processus de défense chez les invertébrés. A l’époque, (1963-1968), l’immunité des vertébrés, avec la présence d’anticorps dont la structure avait été récemment décryptée, régnait en maître. Vouloir s’occuper d’invertébrés, dont les défenses ne répondaient à l’époque à aucun critère exigé en immunologie (cellules de type lymphocyte, mémoire immunitaire, présence d’immunoglobulines) était suicidaire. Il était indispensable, pour être crédible, de s’appuyer sur un diplôme d’immunologie, que j’ai acquis à l’Institut Pasteur.
Toutefois, compte tenu du développement technique à l’époque, il était difficile d’analyser la réponse d’un invertébrés, souvent de petite taille, à une agression parasitaire : seule la technique des greffes chez les invertébrés avait un droit de cité pour les immunologistes.
Une traversée du désert de dix ans s’est donc imposée, où il a fallu rechercher les modèles expérimentaux d’insectes les plus adéquats, en attendant l’avénement des techniques de biochimie et de générique moléculaire, adaptées à la petitesse des organismes retenus.
Vos modèles d’études étaient la drosophile et des hyménoptères parasitoïdes. Pourquoi avoir mené des recherches sur ces insectes en particulier ? Par passion ou par opportunisme ?
Drosophila melanogaster et Leptopilina boulardi (Source : Evolveit)
C’est à partir des années 1980, après avoir beaucoup prospecté les modèles potentiels d’insectes parasites pour l’étude de leur immunité (des prospections nombreuses sur le terrain, en général en milieu tropical, m’ont fait découvrir de nombreuses espèces parasitoïdes d’hyménoptères Cynipidae) que j’ai découvert et décrit une espèce parasite de Drosophila melanogaster, Leptopilina boulardi, spécifique de son hôte, et présentant des populations variées, en particulier du point de vue de leur niveau de virulence. Il devenait alors évident de profiter pleinement de l’outil génétique que procurait D. melanogaster.
Un modèle était né, avec un hôte résistant ou sensible et un parasite virulent ou avirulent. Actuellement, cette espèce d’hyménoptère parasite, dont le génome est séquencé, reste un modèle d’étude pour de nombreuses équipes, françaises ou étrangères.
Quelles ont été vos découvertes majeures et qu’ont-elles apporté comme connaissances ?
Anthony J. Nappi (1937-2014) (Source : lien)
J’ai donc pu, avec un tel modèle, rechercher le déterminisme génétique exact de cette résistance chez l’hôte et de la virulence chez le parasite. La découverte majeure a été de montrer que ces deux caractères avaient un déterminisme monogénique (i.e. un seul gène intervenant dans chacun de ces processus). A l’époque, en 1984, j’ai eu la chance de rencontrer un chercheur américain, le Professeur Anthony J. Nappi, de Loyola University (Chicago), qui travaillait exactement sur ce modèle, mais avec des outils de microbiochimie. Une collaboration est née, qui s’est perpétuée sur plus de vingt ans.
En 1998, une collaboration avec une biologiste moléculaire, le Professeur Marylène Poirié (Université de Nice-Sophia-Antipolis), nous a permis d’accéder à la nature du gène de résistance et à sa localisation chromosomique.
Qu’est-ce qui vous passionne le plus chez les insectes (biologie, écologie, physiologie…) ?
C’est indéniablement leur diversité, tant au niveau des espèces qu’on niveau infra-spécifique, avec des populations génétiquement différenciées, touchant ainsi du doigt leur adaptation à leur environnement, ou, pour les espèces de parasitoïdes, leurs mécanismes variés d’adaptation à l’insecte-hôte.
J’ai eu la chance de travailler au Etats-Unis et de participer à de nombreux congrès, et de nouer ainsi une relation solide, tant sur le plan scientifique qu’humain, avec mon collègue Anthony J. Nappi. En fin de carrière, j’ai voulu savoir quelles avaient pu être les relations entre entomologistes français et américains au cours de l’histoire. Ainsi a germé l’idée de cet ouvrage, pour retracer l’apport réciproque des deux communautés, de part et d’autres de l’Atlantique.
Malgré le potentiel pluridisciplinaires qu’offre l’étude des insectes (alimentaire, biomimétisme, écologique…), le nombre d’entomologistes ne cesse de diminuer en France. Quel est votre sentiment quant au futur de cette discipline ?
Drosophila melanogaster (Source : Andres Moline – Flickr)
Je m’inscris un peu en faux vis à vis de cette assertion. Il faut bien sûr s’entendre sur le terme d’entomologiste, que vous définissez à juste titre, et que j’approuve, comme une personne qui s’intéresse à l’étude des insectes. Dans ce cas, les travaux réalisés en France n’ont jamais été aussi nombreux dans les domaines variés de la biodiversité, de l’écologie, de la génétique et du comportement chez les insectes. Des enseignements existent en France, ainsi que des recherches diversifiées, où l’insecte est choisi pour l’étude des différents aspects de sa biologie.
A chaque interview, Passion-Entomologie demande à l’auteur de nous raconter une anecdote entomologique, quelle est la vôtre ?
Yves Carton (Source : Y. Carton)
Ce ne sera pas à proprement parler une anecdote « entomologique », mais une rencontre humaine. En 1970, au sortir de ma thèse, j’ai rencontré à Washington un américain qui m’a proposé de travailler avec lui. Ne me sentant pas encore assez armé scientifiquement, j’ai décliné son offre. Quinze ans après, je l’ai retrouvé à une réunion à Montpellier où nous avons décidé de mettre en commun nos recherches : ce collègue s’appelait Anthony J. Nappi, et cette amitié s’est poursuivi plus de trente ans…
L’Afrique du Sud est connue pour sa flore extraordinaire et sa faune endémique. Le Muséum du Cap, l’un des plus grands Muséum africains, possède plus d’un million de spécimens d’arthropodes provenant essentiellement du continent africain.
Holotype de Capederces madibai. A : habitus ; B : vue ventrale ; C : détails de la face ; D : vue latérale (Source : Maquart & Van Noort, 2017)
C’est en parcourant ses collections de longicornes (Coléoptères de la famille des Cerambycidae) que Pierre-Olivier Maquart est tombé sur une série de spécimens singuliers qui se sont révélés appartenir à une espèce nouvelle. Ces sept insectes avaient été capturés entre 1926 et 1928 dans la province d’Albany dans le Cap Oriental. Ils appartiennent au genre Capederces qui fut décrit seulement en 1999 par l’entomologiste autrichien Karl Adlbauer. La seule espèce jusqu’alors connue avait été capturée non loin de là, près de Steytlerville : Capederces hauseri (photos en bas de page). Ces petits longicornes floricoles, aux motifs délicats, sont restés très discrets puisqu’aucun autre spécimen ne fut rencontré depuis leur capture il y a près de 90 ans.
Cette découverte s’est faite en 2013, l’année de la mort de Nelson Mandela. Les auteurs ont donc décidé de dédier cette espèce nouvelle au héros Sud-Africain, surnommé localement Madiba : Capedercesmadibai (photos ci-dessus).La description a été publiée le 22 mars 2017 dans la revue Zootaxa (lien).
L’Afrique du sud héberge 853 espèces de Cerambycidae, beaucoup d’autres restent encore à découvrir…
Le mot du co-descripteur de l’espèce :
Qu’est ce qui a retenu votre attention sur ces spécimens ?
Ils étaient rangés dans une boîte dédiée à la tribu des « Clytini » : leur phénotype inhabituel m’a interrogé et empêché de me décider quant à leur positionnement systématique, éveillant ainsi mes soupçons.
Comment avez-vous vérifié que l’espèce n’était pas décrite ?
Pierre-Olivier Maquart dans les collections du Muséum du Cap – Afrique du Sud (Source : Maquart, 2017)
J’ai découvert que les espèces n’appartenaient pas aux Clytini, mais à la tribu des Tillomorphini (un groupe essentiellement représenté en Amérique du Sud), une photo postée sur le forum de Francesco Vitali a permis de mettre un nom de genre sur l’insecte en question : Capederces.
Ce genre, monospécifique jusqu’à cette découverte (qui ne comporte qu’une seule espèce) était représenté en Afrique du Sud uniquement par un seul mâle de C. hauseri. Les spécimens du Muséum n’étant que des femelles, j’ai donc pensé au départ qu’il pouvait s’agir de cette espèce. Puis, la réception d’une photo d’un spécimen femelle de C. hauseri provenant de l’entomologiste américain Larry Bezark – Department of Food and Agriculture, Sacramento – Californie – a confirmé qu’il s’agissait d’une nouvelle espèce.
Pourquoi l’insecte n’a-t-il pas été revu depuis 90 ans ?
Je ne sais pas… Il faudrait étudier et prospecter dans la zone (« Albany district »). D’un côté, j’ai eu accès récemment aux collections de l’Université de Rhodes, qui contient près de 2 000 de Cerambycidae (dont la majorité a été collectée dans les environs de cette région), où C. madibai n’était pas présente. La faune sud-africaine est largement sous-prospectée et sous-étudiée. Pour en donner un exemple, j’ai reçu des spécimens de la famille des Disteniidae de la région du Cap : Phelocalocera queketti, un longicorne relativement « primitif » qui n’a pas été cités dans la littérature depuis près de 60 ans, alors qu’elle est localement fréquente. Qui sait, peut-être retrouvera-t-on Capederces madibai prochainement ?
Description morphologique de Capederces madibai
Cette nouvelle espèce se distingue de C. hauseri par une coloration entièrement noire avec 4 tâches blanches/ivoires clairement délimitées et des antennes dont la longueur est plus petites que celle du corps (ratio : 0,95 pour C. hauseri et 0,7 pour C. madibai) (voir photos ci-dessous).
– La tête : front plat densément et uniformément ponctué ; base des antennes aplatie et largement espacée ; partie inférieure des lobes optiques plus longue que la zone occipitale ; vertex et scape densément et grossièrement ponctués ; scape avec une échancrure longitudinale distale nette ; 3ème antennomère allongé dont la longueur équivalente aux antennomères 4 et 5 réunis ; antennes de couleur brun foncé et partiellement glabre
– Abdomen : tégument noir ; premier sternite abdominal densément recouvert de soies blanchâtre
– Elytres : plus large que longue (ratio : 2/3) ; surface à ponctuation dense et grossière en nid d’abeille, lisse à l’apex et en périphérie, de couleur noire comportant deux tâches bien définies de couleur blanchâtre/ivoire ; peu de variations entre les spécimens
A gauche : Holotype de Capederces madibai. A : habitus ; B : vue ventrale ; C : détails de la face ; D : vue latérale A droite : Capederces hauseri. A : habitus général de la femelle ; B : vue latérale de la femelle ; C : Habitus de l’holotype mâle (Source : Larry Bezark ; Maquart & Van Noort, 2017)
Source :
– Maquart P.O. & Van Noort S. (2017) : Description of a new species of Capederces (Coleoptera : Cerambycidae : Cerambycinae) from South Africa. Zootaxa 4244(4): 583-587 (lien)
– Retrouvez également l’interview de Pierre-Olivier Maquart : lien
Dans l’univers des parasitoïdes (lire cet article), l’ovipositeur, également appelé oviscapte (appendice abdominal avec lequel les oeufs sont déposés), a un rôle crucial puisqu’il permet de situer précisément la ponte dans l’espace et dans le temps, ce qui est essentiel quand celle-ci doit s’accorder avec le cycle biologique de l’hôte. Avant d’aborder certaines particularités étonnantes, il faut d’abord en détailler quelque peu la structure. Chez la plupart des insectes holométaboles, l’ovipositeur est une simple excroissance tubulaire autour de l’orifice génital, mais, chez les hyménoptères, il a conservé une structure primitive bien plus complexe, qualifiée de « lépismoïde » : il s’agit en fait de deux paires de valves creuses qui forment l’ovipositeur à proprement parlé, et sont protégées par une troisième paire, le fourreau.
Illustration 1 : Coupe transversale d’un ovipositeur « standard » d’hyménoptère, montrant les deux paires de valves et leur articulation coulissante. En médaillon, photo MEB montrant les cténidies qui tapissent la face interne de l’oviducte et empêchent le reflux de l’oeuf lors de la ponte (Source : simplifié d’après Quicke, 2015*)
La première paire est coaptée en une pièce supérieure unique qui s’emboite dans une deuxième paire formée de deux valves inférieures, le tout formant un tube autour de l’oviducte depuis l’orifice génital (illustration 1 ci-contre). Ces deux paires sont liées par un système mécanique formant une glissière longitudinale : la face inférieure des valves supérieures forme un ergot qui s’insère dans un rail porté par les valves inférieures. Ainsi, les deux paires peuvent coulisser l’une par rapport à l’autre, ce qui joue un rôle capital lors de l’oviposition. En effet, l’ovipositeur lui-même ne contient pas de muscle : le passage de l’oeuf y est assuré à la fois par le frottement relatif des valves creuses et par la contraction des muscles abdominaux. La face interne des valves est par ailleurs dotée de structures en peignes empêchant le reflux de l’oeuf durant la ponte.
Curieusement, certains ovipositeurs ont des formes particulièrement sinueuses (Photo 1 ci-dessous). On s’est longtemps interrogé sur l’utilité d’une semblable géométrie, a priori plus gênante qu’autre chose lorsqu’il s’agit de perforer un substrat. Il est de tout de même plus facile de perforer le sol avec une aiguille qu’avec un ressort… Puis on a réalisé que le mouvement relatif des valves d’un ovipositeur sinueux a ceci d’intéressant qu’il en modifie l’orientation verticale. Ainsi, en jouant sur le frottement des deux valves, la femelle en guide l’extrémité. Vous saisissez l’utilité du dispositif lorsqu’il s’agit d’atteindre un hôte enfoui dans une galerie non rectiligne ? La recherche médicale s’est d’ailleurs empressée de récupérer cette petite merveille naturelle pour faire naviguer les instruments de microchirurgie à l’intérieur des vaisseaux sanguins.
Photo 1 : Habitus de Pristomerus hansoni – Rousse, Villemant & Seyrig. Notez la grande taille de l’ovipositeur et surtout la forme très sinueuse de son extrémité (Source : photo – Rousse / Iziko SA Museums)
La structure de base de l’ovipositeur est donc similaire chez tous les parasitoïdes. Mais cette unité structurelle est associée à une très forte diversité morphologique : la forme et la taille de l’organe varient énormément d’un groupe à l’autre et à l’intérieur même d’un groupe. La raison en est simple : la structure générale de l’ovipositeur est liée à la phylogénie, mais sa forme précise est conditionnée par l’écologie. Autrement dit, les hyménoptères, ayant tous un ancêtre commun, ont hérité d’un ovipositeur de structure analogue, mais ces hyménoptères ayant des exigences écologiques très variables, leur ovipositeur a évolué secondairement afin de s’adapter aux conditions environnementales.
Le premier trait directement lié à l’écologie du parasitoïde et à son hôte est la longueur de l’ovipositeur. Les parasitoïdes d’hôtes exposés en surface auront un ovipositeur extrêmement court, alors qu’il atteint une longueur relative impressionnante chez les parasitoïdes qui doivent fouiller de grandes profondeurs. Cette longueur relative est souvent limitée à environ 1,3 fois la taille de l’insecte, ce qui correspond grosso modo à la taille qu’il doit avoir pour que l’insecte, en extension sur ses pattes antérieures, puisse en ramener la pointe vers l’avant et diriger sa ponte. Cependant, de nombreux parasitoïdes ont développé des comportements ou des structures anatomiques qui leur permettent de dépasser cette limite. Dans des exemples extrêmes, cet ovipositeur peut ainsi faire plus de dix fois la longueur du corps de l’insecte.
Photo 2 : vues dorsale et latérale d’une femelle d’Ibalia leucospides avec son gastre en lame de couteau à l’intérieur duquel est enroulé l’ovipositeur (Source : B. Mallet, Le monde des Insectes)
Chez la plupart des parasitoïdes, l’ovipositeur est externe et pointe vers l’arrière à l’extrémité de l’abdomen (appelé également gastre). Lorsque cet ovipositeur est long, l’insecte le traine derrière lui, ce qui peut gêner son mouvement et le pénaliser face aux prédateurs. Certaines espèces à ovipositeur de grande taille ont développé au cours de l’évolution des structures de rangement que l’on imagine facilement plus confortables. Par exemple, dans la famille des Cynipidae, l’ovipositeur forme une boucle plus ou moins complète à l’intérieur de l’abdomen (photo 2), et il est complètement enroulé chez les Orussidae, des parasitoïdes de larves xylophages. Chez les Leucospidae (photo 3) et certains Platygstridae (photo 4), l’ovipositeur est dirigé vers l’avant et rangé sur la face dorsale de l’abdomen au repos.
Photo 3 : Femelle Leucospis dorsigera avec le fourreau de l’ovipositeur reposant sur la face dorsale du gastre et pointant vers l’avant (Source : D. Geystor, Le monde des Insectes)
Dans d’autres cas où l’ovipositeur est particulièrement long, le déploiement est facilité par des structures spécifiques. Ainsi, chez les Ichneumonidae du genre Megarhyssa, la partie externe de l’ovipositeur peut atteindre plus de trois fois la longueur du corps. On peut voir sur cette vidéo comment la femelle de Megarhyssa atrata procède pour insérer et retirer son ovipositeur dans un tronc dans lequel elle recherche un hôte (voir également les vidéos ci-dessous) . Cette délicate manoeuvre est assurée par la rotation complète des segments apicaux de l’abdomen couplée au déploiement d’une membrane issue de la jonction entre les segments 7 et 8, membrane elle-même associée à des glandes sécrétrices. Grâce à cet ensemble, M. atrata peut atteindre des hôtes enfouis sous 14 cm de bois dur, alors que le corps de l’animal ne mesure que 4 cm !
Photo 4 : Femelle d’Inostemma boscii avec le premier segment du gastre transformé portant un cornet courbet abritant l’ovipositeur au repos (Source : J. Svabik)
L’extrémité de l’ovipositeur est également un bon indicateur de l’écologie de son propriétaire. Les parasitoïdes ayant pour hôtes des oeufs, immobiles et fragiles, possèdent dans la majeure partie des cas un ovipositeur à l’extrémité effilée, alors qu’un hôte mobile, comme une larve vivante, requiert une extrémité en tête de flèche barbelée afin d’assurer la prise. Plus surprenante encore est la composition chimique de cet ovipositeur. Ainsi, pour les espèces dont les hôtes se trouvent dans un substrat dur comme le bois, l’ovipositeur est alors renforcé par des minéraux métalliques (manganèse ou zinc) qui en assurent la dureté !
Pascal Rousse (Source : Pascal Rousse)
Passion-Entomologie souhaite grandement remercier Pascal Rousse pour sa nouvelle contribution. Pascal est chercheur entomologiste, correspondant du Muséum National d’Histoire Naturelle de Paris (MNHN), versé dans la taxonomie, l’écologie et la phylogénie des hyménoptères parasitoïdes, en particulier dans le cadre de leur utilisation en lutte biologique, auteur d’une trentaine de publication sur ces sujets, mais avant tout fasciné depuis bien longtemps par la biologie des Ichneumonidae sans que la psychanalyse ait pu trouver une explication rationnelle à cette monomanie !
Contribuer au financement d’une mission d’exploration scientifique dans le
Massif du Makay – Madagascar
Voici un an, avril 2016, je découvrais, parcourais et explorais une région à la fois isolée, riche d’une biodiversité unique, demeurant encore relativement préservée des pressions anthropiques et peu connue : le Massif du Makay.
Ce voyage au bout du monde, comme éco-volontaire et entomologiste, avec les équipes de Naturevolution, m’a permis de confirmer l’intérêt scientifique d’étudier plus profondément cette région pour agir en faveur de sa préservation. Durant son exploration, j’ai réalisé un travail de collecte d’insectes (entomofaune) dans le but de participer à l’inventaire de la biodiversité.
Du 22 juillet au 12 août 2017, je participe à la mission d’exploration scientifique d’envergure menée par Evrard Wendenbaum, fondateur de l’association Naturevolution et un groupe d’étudiants du Muséum d’Histoire Naturelle de Paris : La Société des Jeunes Explorateurs.
Vous pouvez également trouver mon témoignage sur le site de Naturevolution.
Objectif de la mission
L’objectif de cette mission est double : 1) réaliser un inventaire de la biodiversité ; 2) récolter des données biologiques et géologiques qui apporteront les connaissances et les preuves scientifiques du caractère exceptionnel et unique de cette région, auprès des autorités malgaches afin que des mesures de préservation et de conservation soient entreprises. Grâce aux précédentes missions menées par Naturevolution, comme celle de 2011 (extrait vidéo ci-dessous), le Massif du Makay fait désormais l’objet d’un statut d’aire protégée.
La mission, pluridisciplinaire, est composée de nombreux spécialistes : botanique, ichtyologique, mammalogie, ornithologie, géologie… et entomologie, dont j’ai la responsabilité (lire ce lien)
Les activités entomologiques consisteront à :
Explorer des sites et des écosystèmes peu ou pas étudiés présentant un fort intérêt scientifique comme la canopée, les mares temporaires et isolées, les souterrains (humides et secs) ou encore les grotte à chauve-souris.
Collecter un certain nombre de spécimens de diverses familles caractéristiques des différents habitats visités. Remarques : les prélèvements seront limités et raisonnés pour préserver l’équilibre et l’intégrité de populations pouvant parfois être très restreintes
Identifier les spécimens et les classer par ordre, famille et sous-famille pour une détermination plus précise auprès de spécialistes au retour de mission (ce travail demandant une expertise et un matériel spécifique, celui-ci ne peut s’effectuer sur place)
Conditionner les collectes : les spécimens seront placés dans des tubes avec de l’alcool pour garantir un bon état de conservation, et référencés dans une base de données indiquant les coordonnées GPS, le nom du collecteur, l’habitat et le jour de la collecte. Remarques : les spécimens de grande taille et facilement identifiables comme les Odonates (libellules) ou les Lépidoptères (papillons) seront étudiés uniquement sur photos afin de limiter les prélèvements
Inventorier et valoriser : l’ensemble des données collectées durant la mission (nombre d’espèces, répartition, taux de diversité, écologie…) va permettre de compléter l’inventaire et la connaissance de l’entomofaune, valorisable scientifiquement : publications, description possible de nouvelles espèces ; enterrées de communication (vidéos, reportages, exposition, articles de vulgarisation…) ; et en démontrant la richesse biologique unique du Massif auprès des autorités malgaches
Pour en savoir davantage sur les méthodologies de collecte des insectes et leur conditionnement : lire ces articles.
Comment contribuer ?
Vous portez un intérêt aux problématiques de préservation de l’environment et de la biodiversité, vous vous demandez comment contribuer à votre échelle : cette mission constitue l’occasion de participer à l’un des plus grands défis et enjeux du 21ème siècle !
Pour cela, vous pouvez effectuer un don de plusieurs manières :
Don défiscalisable (66%) : rendez-vous sur le site de ProjetsPlusActions (pour les personnes payant des impôts en France uniquement)
Don de matériels ou autre : pour cela, me contacter directement à cette adresse : passion-entomologie@hotmail.fr
La mission se déroulera du 22 juillet au 12 août 2017 (3 semaines). L’ensemble des frais s’élèvent à 3 000€.
Ils se distribuent ainsi :
1 950€ : logistique, matériels, déplacement sur place
1 100€ : avion (A/R)
Qui suis-je ?
Passionné depuis toujours par le monde vivant et tout particulièrement par l’entomologie, mon souhait a été dès mon adolescence d’étudier les insectes et d’explorer les écosystèmes les plus riches de la planète. Mon Master Recherche en entomologie obtenu en 2008 à l’Université de Tours (Institut de Recherche sur la Biologie des Insectes – IRBI), j’ai effectué plusieurs missions scientifiques en France et à l’international : CIRAD-La Réunion, MNHN-Paris, Station des Nourgaues-Guyane française, STRI-Panama (pour en savoir davantage, retrouvezmon CV).
Actuellement je suis chargé de Recherche et Développement en entomologie au sein d’une Start-up dont l’objectif est de valoriser par l’insecte les déchets organiques d’origine végétale (appelés bio-déchets) en protéines animales (insectes) destinées à l’alimentation piscicoles.
Je vous remercie de votre contribution, de votre intérêt à ma passion et mon engagement en faveur de la préservation de l’environnement et de sa biodiversité.
Vous serez bien évidemment informés des suites de la mission par la publication régulière de comptes-rendus.
Album photos de la mission 2016
Recommandation d’ouvrage et DVD sur cette thématique
Professeur émérite à l’Université Paul-Valéry Montpellier 3, chercheur au Centre d’Ecologie Fonctionnelle et Evolutive (CEFE). Spécialiste des mouches piqueuses (glossines, stomoxes) et des parasites transmis. Il est co-éditeur du traité Entomologie médicale et vétérinaire, IRD Editions/Quae, 2017.
Pourriez-vous nous dire quelques mots de votre parcours ?
Ancien élève de l’Ecole Normale Supérieure (rue d’Ulm), agrégé des Sciences Naturelles, Professeur émérite à l’Université Paul-Valéry Montpellier 3, je suis chercheur au CEFE (Centre d’Ecologie Fonctionnelle et Evolutive – UMR 5175), et ma spécialité est l’entomologie médicale.
Gérard Duvallet (Source : G. Duvallet)
J’ai passé 21 ans de ma vie – de 1973 à 1994 – en Afrique de l’Ouest, au Burkina Faso, à étudier les glossines (mouche tsé-tsé) (voir photo plus bas dans l’article) et les trypanosomes qu’elles peuvent transmettre (maladie du sommeil chez l’homme et nagana chez l’animal). J’ai été détaché de l’Education Nationale successivement à l’ORSTOM (devenu l’IRD, Institut de Recherche pour le Développement), puis au Ministère de la Coopération, puis au CIRAD (Centre de Coopération Internationale en Recherche Agronomique pour le Développement), avant de revenir dans mon corps d’origine. De retour en France, j’ai lancé des recherches sur les mouches piqueuses peu étudiées (les stomoxes et les taons), des recherches que l’on continue de mener en France, en Thaïlande et en Afrique.
Je participe actuellement à un projet ANR (Agence Nationale de Recherche) intitulé FlyScreen, qui cherche à mettre au point de nouveaux outils de lutte non polluante contre ces mouches piqueuses.
Le dernier « Précis d’entomologie médicale et vétérinaire » de F. Rodhain et C. Perez (éditions Maloine), en français, utilisé par des centaines d’étudiants, datait de 1985. Il n’est plus disponible en libraire et les connaissances dans ce domaine ont beaucoup évolué depuis cette époque. Il était donc devenu nécessaire de proposer un nouvel ouvrage, mis à jour, et qui devrait servir de référence aussi longtemps que le précédent. Cela ne pouvait être réalisé qu’en regroupant les meilleurs spécialistes. C’est pourquoi ce nouvel ouvrage est l’oeuvre d’un collectif de 41 auteurs que j’ai eu l’honneur de co-éditer avec mes collègues Vincent Robert et Didier Fontenille.
Cet ouvrage est divisé en trois parties qui envisagent successivement : 1) les concepts de méthodes utilisées en Entomologie médicale et vétérinaire, 2) l’histoire de l’Entomologie médicale et les questions de santé publique et d’environnement, 3) les arthropodes d’importance médicale ou vétérinaire.
L’entomologie médicale et vétérinaire s’entend au sens large, en englobant non seulement les insectes, mais aussi les autres arthropodes qui peuvent poser des problèmes en termes de santé humaine ou animale (les tiques, d’autres acariens, des crustacés). Et il ne s’agit pas de traiter uniquement d’agents infectieux qui peuvent être transmis par des insectes, mais aussi de toutes les nuisances possibles : arthropodes venimeux, allergisants, urticants, vésicants ou simplement nuisants en cas de pullulation.
Quelles sont les motivations à écrire une telle somme ?
Les motivations étaient multiples. D’abord actualiser un ouvrage ancien, qui a été utilisé pour la formation de nombreux entomologistes médicaux et vétérinaires francophones. Nécessité donc d’avoir un document à jour des dernières connaissances grâce à la participation des meilleurs spécialistes. Ensuite relancer les formations académiques en Entomologie médicale et vétérinaire, à un moment où, en raison des changements globaux, des insectes et des pathogènes parcourent le monde en quelques heures grâce aux moyens de transports modernes.
L’Institut Pasteur de Paris maintient un cours d’Entomologie médicale, et en a même fait cette année un MOOC qui a un énorme succès au niveau mondiale. L’Université de Montpellier, en association avec l’IRD et l’Université Alassane-Ouattara a Bouaké en Côte d’Ivoire, a créé il y a une dizaine d’année un Master International d’Entomologie médicale et vétérinaire qui forme chaque année une dizaine d’entomologistes médicaux venant d’Afrique, d’Europe, d’Amérique du Sud et de l’Océan Indien.
Cet ouvrage, qui sera distribué gratuitement à ces étudiants grâce au mécénat, constituera un appui fort pour ces formations. Il permettra aussi aux anciens étudiants de maintenir leurs connaissance à niveau.
Certaines espèces d’insectes sont vectrices de maladies et de pathogènes. Malgré les progrès techniques et scientifiques du XXème siècle, celles-ci n’ont pu être contenues : quels sont et quels ont été les principaux obstacles et freins aux progrès dans ce domaine ?
Notons d’abord que les vecteurs sont vecteurs d’agents infectieux et non de maladies. La maladie est ensuite un dialogue singulier entre l’hôte et l’agent infectieux inoculé. Le vecteur ne joue un rôle dans la maladie qu’en inoculant l’agent infectieux et aussi, parfois, en favorisant son implantation grâce aux propriétés de sa salive injectée en même temps.
On a cru en effet au XXème siècle que l’on viendrait à bout de ces insectes vecteurs ou nuisants grâce aux insecticides comme le DDT. On a bien vu qu’il n’en était rien. Simplement parce que nous avons affaire ici à des systèmes évolutifs et à des organismes qui ont des capacités énormes d’adaptation. Les insectes sont rapidement devenus résistants aux insecticides. On a beau avoir inventé de nouveaux insecticides, les insectes se sont adaptés. Et les agents pathogènes sont aussi devenus résistants aux médicaments utilisés, comme les bactéries sont devenues résistantes aux antibiotiques. C’est une course permanente. Et une course folle, car, pendant longtemps, nous n’avons pas mesuré les effets collatéraux de ces traitements. Il a fallu que paraisse en 1962 l’ouvrage « Silent Spring » de Rachel Carson pour alerter le monde sur les effets néfastes des insecticides sur les oiseaux et sur la biodiversité en général. Or on sait maintenant le lien fort entre biodiversité et santé, ainsi qu’entre biodiversité et fonctionnement de nos écosystèmes. Il faut repenser le contrôle de ces nuisances dans un cadre écologique global. Et cet ouvrage devrait y aider.
Sommaire de « Entomologie médicale et vétérinaire » (Source : G. Duvallet)
Sommaire de « Entomologie médicale et vétérinaire » (Source : G. Duvallet)
Sommaire de « Entomologie médicale et vétérinaire » (Source : G. Duvallet)
Mal considérés et perçus uniquement comme « nuisibles » par une grande majorité de la population, quels sont les principaux bénéfices que peuvent pourtant apporter les insectes dans les domaines de la santé humaine et animale ?
Il est clair qu’en ne parlant uniquement que des vecteurs ou des arthropodes nuisants, on pourrait transmettre au public une image totalement fausse. Les insectes dans leur immense majorité jouent un rôle très positif. Si l’on pense aux services écosystémiques, on pourrait citer quelques exemples :
– Les insectes pollinisateurs : abeilles domestiques, abeilles sauvages, de nombreux diptères et autres insectes. Sans eux, nous n’aurions pas nombre de fruits ou de légumes sur nos tables ;
– Les insectes coprophages qui incorporent dans le sol les excréments des animaux. Ces bousiers jouent un rôle considérable en favorisant la fertilité des sols et en nettoyant les prairies ;
– Les insectes nécrophages et/ou nécrophiles qui aident à faire disparaître les cadavres ;
– Les parasitoïdeset autres prédateurs qui contrôlent naturellement les populations d’autres insectes : libellules, syrphes, carabes, etc.
Un domaine particulier est celui de l’asticot-thérapie. De nombreux centres existent en Grande Bretagne, pratiquement aucun chez nous. On sait que certaines larves de mouches, des asticots, sont en effet capables de nettoyer des plaies et de favoriser la cicatrisation. Ces observations proviennent de médecins militaires qui travaillaient sur les théâtres de guerre, notamment dans les tranchées au cours de la Première guerre mondiale. L’utilisation de ces asticots peut permettre éviter des amputations dans des cas de gangrènes.
Un autre domaine particulier est celui de l’utilisation des insectes dans l’alimentation. De nombreuses populations en Afrique, en Asie ou en Amérique du Sud, incorporent les insectes à leur alimentation. C’est clairement un domaine d’avenir en raison des nombreux avantages de ces apports.
Le nombre d’espèces d’insectes actuellement connues est environ de 1,3 millions. Et les entomologistes estiment qu’il en existe environ 8 millions d’espèces. On est donc loin d’avoir encore tout inventorié. Et les vecteurs et les nuisants ne sont qu’une infime partie de cette énorme biodiversité. Il faut faire en sorte que les méthodes de lutte contre ces vecteurs n’impactent pas le restent de la biodiversité et il faut soutenir les recherches dans ce domaine.
Les stratégies et les solutions de lutte doivent être adaptées pour être plus efficaces et protectrices de l’environnement et de la santé humaine, comment ce changement peut-il et doit-il s’opérer dans les années à venir ?
Accouplement de Glossina morsitans – Mouche tsé-tsé ou mouche du sommeil – L’absorption de sang par la femelle est visible à travers son abdomen (Source : Ray Wilson)
On a pris beaucoup de retard, en pensant que les insecticides résoudraient tous ces problèmes. Heureusement, des recherches se sont développées qui analysent le fonctionnement des écosystèmes et mesurent plus clairement les impacts. Elles doivent permettre de mieux comprendre ce qui facilite un bon équilibre et limite les pullulations de nuisants. L’écologie chimique peut permettre aussi de comprendre ce qui attire ou ce qui repousse spécifiquement ces insectes et arthropodes nuisants. Cela devrait déboucher sur de nouvelles méthodes de contrôle. Il ne s’agit plus maintenant de pulvériser des insecticides dans l’environnement, mais d’attirer spécifiquement les arthropodes cibles dans des pièges où l’on pourra les contrôler sans nuire au reste de l’environnement. Là encore, de nombreuses recherches restent indispensables.
Etes-vous optimiste quant au contrôle et à l’éradication des nuisances et maladies dues aux insectes ? Notamment dans les pays du Sud ?
L’expérience passée a appris aux entomologistes de ne plus parler d’éradication mais bien de contrôle. Il faut baisser le niveau d’abondance de certaines populations d’arthropodes en dessous de leur niveau de nuisance. Si des moyens sont donnés au pays du Nord comme du Sud pour renforcer la formation des entomologistes médicaux et vétérinaires, et pour soutenir la recherche dans les domaines évoqués (en particulier en écologie fonctionnelle en écologie chimique, en écophysiologie) et la pluridisciplinarité dans ces activités sur le terrain, on pourra être optimiste. Des maladies à transmission vectorielle comme la filariose lymphatique, comme la trypanosome humaine africaine (maladie du sommeil), et d’autres sont en diminution au regard des chiffres de l’OMS. Les recrudescences interviennent dans le cas de conflits armés ou de guerres civiles lorsque médecins et épidémiologistes ne peuvent plus aller sur le terrain dépister et traiter les porteurs de pathogènes.
Les mantes, groupe fascinant d’insectes qui sont connus de tous pour leur voracité (certains diront leur cannibalisme), sont visibles quasiment partout dans le monde. Seuls les pôles et les milieux d’altitude très élevée n’accueillent pas d’espèces. Ailleurs, elles se rencontrent des déserts africains aux zones humides et relativement vierges des différents grands blocs forestiers de la planète, en passant par les régions tempérées, les montagnes…
Couple de Mantis religiosa – Mâle étant sur la femelle, plus grande (Source : N. Moulin)
Les mantes sont les plus diversifiées en Afrique (plus de 900 espèces décrites), puis en Asie (plus de 550 espèces), en Amérique (plus de 420 espèces), en Océanie (plus de 165 espèces), enfin en Europe (avec environ 25 espèces). En France métropolitaine, il est possible d’en observer 8 espèces, dont la plus connue est Mantis religiosa (Mantidae), visible du sud au nord de l’Hexagone (photo ci-contre).
Ressemblante, mais beaucoup plus méridionale, Iris oratoria (famille des Tarachodidae) est remarquable avec ses patterns de couleurs sur les ailes postérieures. Complètement différente, Empusa pennata (Empusidae), aussi appelé le « diablotin », constitue souvent une figure emblématique dans les ouvrages et revues de nature avec son cône céphalique, ses longues pattes et ses multiples extensions foliacées situées un peu partout sur son corps.
Les autres mantes que l’on rencontre en France, de taille plus réduite, sont souvent géophages ou apprécient la strate herbacée rase. Il y a les Ameles (Mantidae) (Ameles spallanziana et A. decolor) qui se distinguent par leur morphologie plus ou moins trapue et la forme des yeux. Geomantis larvoides (Mantidae), aptère, petite, a un tubercule derrière les yeux. Pseudoyersinia brevipennis (Mantidae) n’a pas été revue dans le Var depuis 1860, alors qu’elle est présente sur le pourtour méditerranéen. Enfin, Perlamantis alliberti, une toute petite mante brune au pronotum presque carré, appartient à la famille des Amorphoscelidae.
Mantes de France
Description des espèces françaises de mantes – Habitus des différentes espèces en bas de page (Source : Nicolas Moulin)
L’ordre des Mantodea appartient au Super-ordre des Dictyoptera, au même titre que les blattes (ordre des Blattodea) (lire cet article). Actuellement, les mantes comptent près de 2500 espèces valides, parmi 16 familles (sans compter les familles des mantes fossiles) : Mantodea.speciesfile.org. Il y a encore 90 ans nous en étions qu’à 1800 espèces environ. Le nombre d’espèce décrites depuis un siècle est dû au travail de « fourmis » de plusieurs entomologistes reconnus mondialement : Beier, Giglio-Tos, Rehn, Kaltenbach, Roy, … Et plus récemment, Svenson, Stiewe, Rivera, … et toujours Roy ! Depuis 1927, date de la dernière grande « faune » sur les mantes, ce ne sont que descriptions d’espèces éparses et révisions de genre qui sont produites. Est-ce qu’un jour, un ou plusieurs entomologistes se chargeront de l’actualiser ? Pourquoi faudrait-il actualiser ?
Depuis 90 ans donc, les publications sur la taxonomie des mantes sont éparses : les descriptions sont isolées dans des articles scientifiques ici ou là ; des révisions de genre, de sous-famille sont publiées, ici en français, là-bas en anglais… Le séquençage ADN (cf. encadré ci-dessous) a fait son apparition. Les études génomiques réorganisent le phylum des Mantodea, avec la création récente, par exemple, de la famille des Galinthiadidae. Si nous souhaitons travailler sur une espèce découverte ou sur un genre, il faut alors collecter toute cette bibliographie pour rassembler les informations recherchées. C’est fastidieux. C’est pour cela, et à l’image d’ouvrages existant sur d’autres ordres d’insectes, qu’il serait nécessaire qu’une clé des mantes soit mise à jour. La taxonomie est continuellement en évolution mais cela permettrait du moins de rassembler les connaissances… Au moins une fois par siècle !
Des progrès taxonomiques
Codes-barres ADN
L’identification des espèces sur les critères morphologiques commence à connaître ses limites. En effet, elle demande un niveau d’expertise important. Les clés de détermination sont éparses et parfois obsolètes. Il existe des espèces critiques, des espèces aux morphologies indifférenciables… C’est pour cela que, depuis 2003, grâce à Paul Hebert et ses collaborateurs, la distinction entre des espèces peut se faire par le biais des codes-barres ADN. Le support utilisé est l’ADN du gène mitochondrial Cytochrome Oxydase-I (COI) car ses amorces sont universelles, robustes et la vitesse d’évolution permet une bonne discrimination entre les espèces. Concernant les mantes, actuellement, mon entreprise gère plusieurs projets dont les spécimens séquencés proviennent des écoles de terrain ECOTROP en Afrique et de ma collection de références. Pour cette dernière, 551 spécimens ont été séquences, représentant 143 espèces.
Toujours est-il que ce groupe d’insectes est en perpétuelle évolution de nos jours. De nouvelles espèces sont régulièrement décrites, souvent à la suite de la découverte d’un seul individu comme, par exemple, Ovalimantis maculata (Acanthopidae) découverte en Guyane française en septembre 2014 à la Montagne des chevaux par la Société Entomologique Antilles-Guyane (SEAG), puis décrite en 2015 par Roger Roy du Muséum National d’Histoire Naturelle (MNHN) dont la description a permis de créer un nouveau genre sud-américain de mantes : Ovalimantis. Ce genre monospécifique n’est donc connu que d’un seul mâle !
Actuellement, grâce à de nouvelles techniques de génomiques, l’identification morphologique classique des mantes est couplée avec le séquençage ADN (barcoding DNA). A cela s’ajoute des études et des thèses sur les traits de vie (morphométrie : taille des ailes, armatures des pattes ravisseuses…). Ces travaux menés par de jeunes entomologistes (principalement américains, allemands et anglais) permettent de dynamiser l’étude taxinomique des mantes.
Par exemple : la thèse de Franck Wieland (Hambourg) a donné une publication sur le système phylogénétique des mantes basé sur leur caractère morphologique, les travaux de taxonomie et le poste d’éditeur sur les Mantodea dans Zootaxa de Julio Rivera (Toronto), les travaux de taxonomie de Martin Stiewe (Londres) et enfin, la coordination du site de références : mantodearesearch.com et les différents travaux de génomiques et taxonomie de Gavin Svenson (Cleveland) (Bibliographie en bas de l’article).
De redoutables prédateurs
Mantes mimétique du genre Haania sp. – Preah Monivong National Park – Cambodge (Source : Paul Bertner)
Revenons à leurs moeurs délicates… Les mantes sont des prédateurs s’alimentant d’insectes. Selon leur morphologie globale ou leur stade de développement, elles se nourrissent soit de petits insectes (collemboles), d’insectes volants (mouches, papillons), ou plus massifs (criquets, blattes). Tout au long de leur développement, les mantes vivent isolées les unes des autres car les rencontres sont souvent fatales pour la plupart des espèces.
Chez certaines, aux armatures des pattes ravisseuses frêles, le cannibalisme ne se rencontre pas. Une fois à l’état adulte, la reproduction est l’objectif principal. Cependant, comme chez la plupart des araignées, la voracité et/ou le dimorphisme sexuel n’est pas pour faciliter les affaires des mâles. Ils risquent à tout moment (avant, pendant, après) de se faire dévorer vivant. Les mâles peuvent être considérés comme des proies et pourraient apporter un supplément nutritionnel aux femelles. A part chez des espèces « primitives » (aux nombres importants de caractères morphologiques uniques et à l’origine de caractères plus évolués, illustrant une plésiomorphie : caractère ancestral) comme celles du genre Metallyticus, où les couples s’apparient tête/bêche à l’image des punaises, les mâles grimpent sur le dos des femelles pour s’accoupler. Ils introduisent un stock de leur semence dans un organe récepteur interne de l’abdomen des femelles, qui pourront alors produire plusieurs oothèques après un seul accouplement. L’accouplement peut démarrer peut démarrer de quelques minutes à plusieurs heures après le positionnement du mâle sur le dos de la femelle. Ensuite, les durées d’accouplement varient selon les espèces : chez Iris oratoria, cela peut durer entre 45 et 110 minutes ; la durée la plus longue enregistrée est de 40 heures !
Empusa pennata femelle (Source : B. GILLES)
Les oeufs pondus en une seule fois dans les oothèques (membrane rigide constituée de protéines entourant et protégeant les pontes de certaines espèces d’insectes) éclosent selon des laps de temps propres à chaque espèce, et variables selon les conditions météorologiques. Par exemple, chez Mantis religiosa, en milieu tempéré, les oothèques passent l’hiver dans la végétation avant que les jeunes ne sortent au milieu du printemps. A l’inverse, en milieu équatorial, les générations se succèdent en continu, selon la durée de développement des embryons dans les oothèques qui peut durer parfois plusieurs mois. Il existe des exceptions (au moins pour les espèces dont nous connaissons la biologie) : Empusa pennata, en France métropolitaine, passe l’hiver sous forme de jeunes plus ou moins actifs selon les températures (photo ci-contre). Les adultes apparaissent au début de l’été, pondent, et les jeunes, nés fin août, réaliseront 2 à 4 mues avant de se stabiliser pour passer l’hiver. Globalement, ce sont quelques jeunes à plusieurs centaines qui effectueront leur première mue en émergeant de l’oothèque, sécheront puis se disperseront dans la nature. Etant hétérométaboles, les mantes, au fur et à mesure des mues (entre 5 et 10 selon les espèces et le sexe), vont grandir. Les organes du vol (s’ils existent) et les organes reproducteurs se développeront pour être fonctionnels après la mue imaginale (dernière mue aboutissant au stade adulte).
La mante orchidée : Hymenopus coronatus
Couple Hymenopus coronatus (Source : N. Moulin)
Hymenopus coronatus, de la famille des Hymenopodidae (Svenson et al., 2015), se rencontre dans la péninsule indo-malaisienne. Cette mante, hors norme, mime avec perfection les inflorescences d’orchidées. La coloration générale blanche, complétée de brun, beige, vert et même rose la confond efficacement avec les fleurs. De plus, des yeux en pointes et des expansions foliacées très développées sur les pattes ajoutent à son camouflage. Le mimétisme se poursuit avec un déplacement saccadé les faisant passer pour des fleurs frémissant au vent. Le dimorphisme sexuel est aussi à noter : les mâles sont près de 4 à 5 fois moins massifs que les femelles. Ces dernières sont donc généralement sédentaires à leur support alors que les mâles, petits et agiles, volent pour les trouver. A la naissance, les jeunes n’arborent pas les couleurs végétales des stades supérieurs : ils ressemblent à des fourmis, à l’image des jeunes d’Extatosoma tiaratum (phasme à tiare), ils ont la tête noire et le corps rouge intense.
L’écologie des mantes est très variable. Si on en revient aux traits d’histoire de vie, leurs habitats sont tout aussi diversifiés que leur morphologie. Comme les phasmes, le mimétisme est le maître mot chez de nombreuses espèces : certaines, fines et élancées, vivent dans les strates herbacées ; d’autres, très plates ou proche de la forme et de la coloration du lichen (photo en début de paragraphe), vivent sur les troncs et branches d’arbres ; certaines, massives, vertes, se distinguent difficilement dans les strates arbustives et arborées ; d’autres ressemblent à des feuilles mortes et se déplacent souvent dans les milieux de branches mortes ou au sol ; enfin, les mantes dites « fleurs » miment la coloration, jusqu’à la forme, des inflorescences de toutes sortes (cf. encadré ci-contre sur la mante orchidée,Hymenopus coronatus).
La diversité morphologique des mantes n’a d’égal que celle des phasmes. Ces deux groupes d’insectes peuvent être champions en taille et en qualité du camouflage. La plus petite espèce connue dans le monde est Mantoida tenuis (Mantoididae), mesurant seulement un centimètre ! Tandis que la plus grande espèce est probablement Ischnomantis gigas (Mantidae), ayant près de 17 centimètre, et évoluant dans les hautes herbes des savanes de l’Afrique de l’ouest. Les espèces les plus massives (en considérant les femelles, car très souvent plus grosses que les mâles) se rencontrent en Amérique du Sud avec le genre Macromantis (Mantidae) et en Afrique avec le genre Plistospilota (Mantidae).
Morphologie des mantes
De manière synthétique, les mantes ont une tête généralement plus large que longue, dans une position orthognathe (perpendiculaire à l’axe du corps). Le vertex et le front peuvent être simples ou ornés de tubercules, pointes, expansions diverses. Les yeux composés sont développés. Ils sont complétés par des yeux simples, ocelles, situés entre les antennes. Ces dernières sont bien développées chez toutes les espèces mais peuvent varier de forme, de l’antenne filiforme aux antennes pectinées (à l’image des papillons de nuit). Les pièces buccales sont de types broyeur.
Le thorax est de forme diversifiée ; c’est ce qui oriente la silhouette générale de la mante : trapue, longiligne, large, avec des expansions foliacées, des tubercules, des pointes… Il porte les trois paires de pattes et les deux paires d’ailes (quand elles sont développées).
Habitus général d’une mante (Source : d’après Ehrmann 2002)
Les pattes antérieures (dites ravisseuses) sont plus ou moins développées et armées d’épines. C’est un point morphologique très distinctif pour différencier les espèces. Le nombre d’épines et leur disposition sont important. Les pattes médianes et postérieures sont quant à elles généralement plus simples, sauf lorsqu’elles sont ornées d’expansions ou lobes comme chez les Toxoderidae, Acanthopidae, Hymenopodidae…
L’abdomen de forme variable est globalement allongé avec dix tergites (dorsaux), neuf sternites (ventraux) chez les mâles et sept chez les femelles. Le dixième (et dernier) tergite constitue la plaque supra-anale, qui est de forme variable également selon les espèces. L’abdomen est terminé par une paire de cerques segmentés. Là aussi, les formes varient, et en particulier celle du dernier segment : il peut être aplati, cylindrique, foliacé. Chez les mâles, le dernier (neuvième) sternite (plaque sous-génitale) porte les styles, très souvent simples de conformation. Dans la concavité de cette plaque se situe les genitalia (organes reproducteurs). Ils sont toujours de forme asymétrique est divisé en trois parties : un hypophallus ventral, surmonté d’un épiphallus gauche et d’un épiphallus droit. Les genitalia des mâles constituent un critère majeur pour l’identification spécifique des mantes… C’est l’histoire de la clé et de la serrure…
Présentation de l’auteur : Nicolas Moulin
Nicolas Moulin à la recherche de mantes dans la canopée – Expédition Sangha – RCA – 2010 (Source : N. Moulin)
Entomologiste freelance, j’interviens depuis plus de dix ans pour des inventaires dans le cadre d’évaluations environnementales, pour des suivis de populations d’insectes, pour des suivis d’espèces protégées. J’effectue également des animations pédagogiques, des formations à l’entomologie, à ,la création d’entreprise. Mes contrats me font voyager dans tout l’Hexagone (dans la limite de mon domaine biogéographique de plaine car je ne suis pas spécialisé dans les espèces d’altitude ou du Bassin méditerranéen) mais aussi dans les DOM-TOM. Ponctuellement, je quitte la France pour l’Afrique principalement, afin d’encadrer des étudiants dans le cadre d’écoles de terrain sous les tropiques (ECOTROP).
Mon travail sur les mantes est non lucratif, mais m’apporte une satisfaction personnelle d’amélioration des connaissances sur la taxonomie et l’écologie de ces insectes fascinants. Je rédige des articles de vulgarisation, de systématique, de biogéographie ou écologie, et je participe à l’enrichissement des banques de séquences d’ADN les concernants.
Habitus des différentes espèces françaises de mantes
Photos d’espèces de mantes
Recommandations de liens internet sur les mantes
– Mantodea.speciesfile.org : Site de recensement de toutes les espèces de mantes avec des données sur la phylogénie, les références bibliographiques, la répartition…
– Mantodearesearch.com : Site de recensement des personnes travaillant sur les mantes, actualités…
– Hebert P.D.N. et al., (2003) : Biological identifications through DNA barcodes. Proceedings of the Royal Society Biological Sciences, 720:313-321 (lien)
– Hebert P.D.N. ; Ratnasingham S. & deWaard J.R. (2003) : Barcoding animal Life : cytochrome c oxydase subunit 1 divergences among closely related species. Proceedings of the Royal Society Biological Sciences, 270:596-599 (lien)
– Rivera J. & Svenson G.J. (2016) : The neotropical « polymorphic earless praying mantises » – Part I : molecular phylogeny and revised higher-level systematics (Insecta: Mantodea, Acanthopoida). Systematic Entomology, 41:607-649 (lien)
– Svenson G.J. et al., (2015) : Of flowers and twigs: phylogenetic revision of the plant-mimicking praying mantises (Mantodea: Empusidae and Hymenopodidae) with a new suprageneric classification. Systematic Entomology (lien)
– Wieland F. (2013) : The phylogenetic system of Mantodea (Insecta: Dictyoptera). Species, phylogeny & Evolution, 3,1:3-222 (lien)
Du 22 juillet au 2 septembre est organisée par Naturevolution une grande expédition scientifique pluridisciplinaire dans le massif du Makay. Cette mission fait suite à la première traversée de cette région par Evrard Wendenbaum en 2007 et aux missions scientifiques de 2010 et 2011.
Le travail de Naturevolution a ainsi permis la création de la Nouvelle Aire Protégée du Makay (pour en savoir davantage : ici).
Présentation de la mission
Les objectifs de la mission sont multiples :
Etudier
Protéger
Valoriser le Makay
Pour cela, une équipe constituée de scientifiques, d’étudiants, d’artistes, d’éco-volontaires et d’acteurs de terrain, de la science et des médias oeuvrera pour étudier, protéger et valoriser ce massif unique au monde.
A l’origine de cette mission et de la démarche scientifique, 5 étudiants du Muséum National d’Histoire Naturelle de Paris se sont réunis en créant la Société des Jeunes Aventuriers dans le but de permettre à de jeunes scientifiques de mener des travaux de recherche sur le terrain et de partager leur passion.
Equipe organisation
Gestionnaire de l’Aire protégée, l’équipe de l’association Naturevolution organise la mission : partenaires, logistiques et projets scientifiques.
Evrard Wendenbaum : Explorateur, fondateur et président de Naturevolution
Philippe Mistral : Responsable mission de terrain – Naturevolution
Bernard Forgeau : Responsable logistique – Naturevolution
Gaëtan Deltour : Responsable scientifique, Secrétaire de Naturevolution
L’expédition a également une vocation pédagogique sur la protection de l’environnement, la sauvegarde de la biodiversité et l’étude de la faune et la flore. Il s’agit de sensibiliser le plus grand nombre de l’intérêt de mener une telle mission et du caractère unique et exceptionnel du massif du Makay.
Pour cela, une équipe de tournage, de photographes, d’illustratrices, de journalistes et de médiateurs scientifiques vont accompagner et assurer le suivi de l’expédition aux côtés des scientifiques et des acteurs de terrain.
Parmi eux, on retrouvera :
Natexplorers : un duo de biologistes, Julien Chapuis et Barbara Réthoré, forts de leurs explorations des forêts d’Amérique Centrale et de leurs talents en médiation scientifique
DirtyBiology : le youtubeur Léo Grasset aux 460 000 abonnés
Il sera possible de suivre le déroulement et les actualités de l’expédition en temps réel à l’aide d’une grande diversité de canaux de communication.
Par exemple, un journal de bord racontera le quotidien en direct du Makay viaFacebook, Twitter et Instagram. La participation d’une équipe de tournage a pour souhait de développer un documentaire long-métrage , une web-série et des vidéos.
L’ensemble des photos, vidéos, illustrations et sons collectés durant l’expédition feront l’objet d’expositions interactives destinées à de grandes structures de médiation scientifique.
Equipe scientifique
Cette mission scientifique est la première à se dérouler en période sèche. Bien que la saison humide soit plus favorable à l’observation, la saison sèche offre la possibilité d’étudier la faune et la flore durant cette période de transition.
Aussi, l’expédition s’intéressera pour la première fois à l’exploration et à l’étude de deux nouvelles zones forestières : Makaykei au sud et Mahasoa au nord.
Les camps de base de la mission 2017 (Source : Naturevolution)
Les projets scientifiques aborderont de multiples champs disciplinaires :
Carnivores, Rongeurs et Chauves-souris : Margot Michaud, Rohan Mansuit, Paul Nicolas (Naturevolution) ; Martin Raheriarisena et Steeve Goodman (Vahatra)
Fougères, Bryophytes, Angiospermes et Canarium : Catherine REEb (Naturevolution) et Rado Ramahandrison (Université d’Antananarivo)
Oiseaux : Vincent Roméra (Agence VISU), Anne-Sophie Lafuite (CNRS) et Eric Temba (Université d’Antananarivo/Asity)
Insectes et Crustacés : Benoît Gilles (Naturevolution) et Jean-François Cart (IUCN)
Archéologie : Antoine Heurtel (Naturevolution)
Poissons : Megann Texereau (Naturevolution) et Jean Robertin Rasoloariniaina (CNRE)
En plus de visiter cette région isolée et unique, l’expédition offre pour la première fois à des éco-volontaires l’opportunité de participer et de contribuer à des projets de recherche scientifique dans un environnement isolé et unique.
Le soutien et l’aide des éco-volontaires permettront de réaliser des échantillonnages d’insectes, des suivis et des comptages de lémuriens, des inventaires botaniques sur les parois verticales des canyons ou encore de visiter des sites archéologiques.
Retrouvez les comptes-rendus de ma mission comme éco-volontaire d’avril 2016 :
J’ai eu le plaisir de m’investir cet été dans une mission d’exploration scientifique pluridisciplinaire dans le massif du Makay à Madagascar (21 juillet – 2 septembre 2017) portée par l’association Naturevolution et Evrard Wendenbaum, son président fondateur.
Mon premier séjour dans cette région, en avril 2016, m’a permis d’approcher le potentiel unique et le potentiel tant faunistique que floristique du massif du Makay. Fort de ce constat, j’ai souhaité m’impliquer et contribuer à la fois à son étude scientifique et à sa préservation en participant à cette mission.
Mon souhait était de répondre à plusieurs objectifs. Il s’agissait de collecter des insectes par l’utilisation de multiples stratégies d’échantillonnage : pièges Malaise et d’interception, filer fauchoir, battage (parapluie japonais), pièges lumineux et attractifs (lire cet article) et de réaliser des inventaires n’ayant jamais été entrepris dans le massif du Makay (inventaires verticaux sur les parois des falaises, inventaires dans la canopée, etc.) afin d’apporter des données nouvelles pour la science et pour argumenter en faveur de la préservation du site.
Une grande quantité de matériel a pu être rassemblée, provenant pour l’essentiel de prêts d’entomologistes à travers la France, tant amateurs que professionnels (liste et remerciement en bas de page).
Déroulement
Situation géographique des camps de l’expédition Makay 2017 (Source : Naturevolution)
Il était initialement prévu au départ de séjourner dans le sud du Makay (voir cet article) (camp Makaikely) mais des changements dans l’organisation sont venus dès le début de la mission contrarier le programme scientifique et logistique. Au lieu de trois camps (nord, centre, sud), ce furent seulement deux camps, au nord, qui furent établis pour les trois premières semaines. Après une semaine de transit, je me suis donc installé pour huit jours dans le camp de Mahasoa, au nord du massif, avec d’autres membres de l’équipe, notamment la botaniste Catherine Reeb (Laboratoire ISYEB-MNHN) et le carcinologiste Jean-François Cart (Expert IUCN).
Le campement se situait dans les forêts de Mahasoa et de Beroha, sur une rive sablonneuse proche de falaises et de forêts à la fois préservées et anthropisées (photos ci-dessus).
Une succession de problèmes d’organisation et de logistique ont perturbé voire empêché un grand nombre d’activités de recherche : changement de programme et prise de décision sans cesse remise en cause, nombre d’écovolontaires trop important et ne permettant pas le transport dans un délai et des conditions raisonnables, un temps de présence sur les camps de ce fait trop court, une réception tardive, voire absente, absence de matériel scientifique et de travail : pas de dôme scientifique, pas de chaises, pas d’électricité en quantité suffisante, pas de table où installer les laboratoires de travail (loupes binoculaires, ordinateurs, cahiers, échantillons, prise de vue photographique…), etc.
Il fut donc plus que regrettable qu’un tel manque d’organisation ait pu venir perturber et empêcher le travail dans une région aussi biologiquement riche et dont l’intérêt était précisément son isolement.
Activités de recherche
Malgré toutes ces difficultés, il a été possible d’installer et d’utiliser les pièges Malaise et lumineux (photos ci-dessous). Afin d’étudier les variations de la diversité et la répartition de l’entomofaune entre différents types d’habitats, les trois pièges Malaise ont été installés dans trois habitats différents : une rive végétalisée, une rive faiblement végétalisme et un sous bois de forêts préservées. Le piège lumineux a été placé près du camp, une fois sur le sable et une autre fois à la cime d’un arbre. Ces collectes ont fait l’objet de dessins par Aurélie Auka (la dessinatrice de la mission) et de mon interview par Léo Grasset (Dirtybiology – Youtubeur) lors du piège lumineux de la canopée (Photos ci-dessous, le dessin d’Aurélie Auka sera prochainement publié et l’interview de Léo Grasset (DirtyBiology) est en attente de publication).
Les inventaires verticaux, quant à eux, n’ont pu être réalisés du fait de l’absence du matériel du moins durant ma présence sur le camp. Cela est décevant car la caractérisation de la répartition de l’entomofaune en fonction de la hauteur, de la luminosité, de l’humidité et de la végétation offrait de belles perspectives d’études scientifiques.
La saison sèche et le froid nocturne expliquent que les collectes aient été peu diversifiées et peu abondantes. Les meilleurs échantillonnages ont été réalisés au filet fauchoir et au piège Malaise, bien que la durée de pose ait été trop courte pour ces derniers (8 jours au lieu de 3 semaines). L’ensemble des principaux ordres (Diptera, Coleoptera, Lepidoptera, Odonata, Heteroptera…) ont été observés, mais la quasi-totalité des captures concernait de petites espèces faiblement représentées.
Premiers résultats
Un premier inventaire a été réalisé sur place : identification, classement et conditionnement par ordre et famille lorsque cela été rendu possible sans matériel d’observation adapté (loupe, lumière, table…). Les individus ont été conditionnés dans l’alcool avec un code référence (lire cet article).
Nombre total d’individus identifiés et classés : 261
Pourcentage Coleoptera : 23%
Pourcentage Heteroptera : 20%
Pourcentage Diptera : 18%
Pourcentage Hymenoptera : 14%
Pourcentage Lepidoptera : 6,5%
Pourcentage Odonata : 3%
Pourcentage Orthoptera : 1,5%
Pourcentage autres : 11%
Pourcentage des captures en fonction des types de pièges
Filet fauchoir et battage : 44,4%
Pièges lumineux : 33,7%
Pièges Malaise : 15% (par manque de temps, les échantillons des pièges n’ont pu tous être identifiés et conditionnés individuellement – la proportion des collectes par ce procédé est en réalité bien plus élevé)
Un inventaire plus complet et exhaustif sera publié dans les mois à venir. Les échantillons, restés à Madagascar dans l’attente des permis d’exportation, seront transmis aux entomologistes spécialistes pour détermination dès leur réception en France.
Des observations naturalistes intéressantes ont été faites par les groupes d’autres disciplines comme l’ornithologie, l’archéologie, l’herpétologie, la mammalogie (lire l’encadré ci-dessous).
Bilan scientifique général de l’expédition
Premières images de fossa (Cryptoprocta ferox) dans le massif du Makay – Plus grand prédateur de Madagascar
Premières découvertes de crustacés d’eau douce (crabes et crevettes) dans le massif du Makay
Observations d’une dizaine de nouvelles espèces d’amphibiens et de reptiles pour le massif, dont le serpent Langaha madagascariensis
Recensement de 10 des 14 espèces de rapaces diurnes de Madagascar
Echantillonnage de plusieurs spécimens de poissons du genre Pachypanchax – Probablement une nouvelle espèce pour la science (une seule population recensée)
Découverte de cinq nouveaux sites archéologique présentant des peintures pariétales
Constat général
Les trois semaines de mission ont été bien trop courtes. Sur 21 jours, la période sur le camp n’a représenté que hui jours. Le repérage du site et l’installation du matériel, comme la pose des pièges, occupent la première journée, puis le rangement de l’ensemble des affaires le dernier. Le temps effectif réservé aux collectes et aux études n’était au final que de six jours : ce qui est insuffisant pour satisfaire les ambitions et les attentes scientifiques.
Projets futurs
De nouveaux projets dans le Makay sont déjà à l’étude et en préparation avec Catherine Reeb et Gaetan Deltour (responsable scientifique de Naturevolution). Avec mes collègues, je suis à l’initiative d’un projet de mise en place et d’établissement d’une station de recherche pluridisciplinaire permanente au coeur du massif du Makay, projet pour lequel j’apporte tout mon soutien, mon temps et mes compétences. Prévu pour 2019, le camp, conçu de la manière la plus écologique possible, offrira les conditions nécessaires et indispensables de confort pour le travail en milieu tropical et isolé à des équipes scientifiques internationales (professionnels et étudiants), mais aussi à un nombre réduit d’éco-volontaires privilégiés, qui se succéderont par rotations successives. Il sera ainsi possible de mener des travaux de recherche aux protocoles plus complets et précis (en biologie, en géologie, en archéologie, etc.) pouvant s’étaler sur plusieurs mois (Gaetan Deltour nous en dit plus dans son interview).
Interview de Gaëtan Deltour
Responsable scientifique de Naturevolution
Gaëtan, peux-tu te présenter en quelques lignes ?
Portrait de Gaëtan Deltour (Source : G. Deltour)
Originaire de Cahors, dans le sud-ouest de la France, j’y ai passé mon enfance entre bois et rivières. Après des études à Toulouse en Licence BOPE « Biologie des Organismes des Populations et des Ecosystèmes », j’ai rejoint le Muséum d’Histoire Naturelle de Paris (MNHN) pour un Master en spécialité SEP « Systématique, Evolution et Paléontologie ». Durant ce parcours, j’ai eu l’occasion de réaliser des stages dans différentes structures : zoo, cabinet vétérinaire, associations qui m’ont permis de développer un réel intérêt pour la conservation en général et la préservation des Lémuriens de Madagascar. Aujourd’hui, je travaille pour l’association Naturevolution en tant que responsable scientifique des différentes missions de l’association.
Comment as-tu rencontré les membres de Naturevolution et quelle est ta mission ?
Ma rencontre avec Naturevolution et ses membres remonte à mes 19 ans. A cette période, je suis en deuxième année de Licence et je dois trouver un stage pour ma troisième année. Je décide alors de réaliser une mission d’écovolontaire à Madagascar et découvre l’association Naturevolution qui travaille autour du Massif du Makay. J’ai donc participé à une de leur mission où j’ai eu l’occasion de fêter mes 20 ans dans un des plus bel endroit du monde ! Après ce séjour, je suis resté en contact avec l’association et c’est deux ans plus tard qu’à l’occasion de l’une de leur assemblée générale, je suis devenu secrétaire de l’association. Depuis, je m’investi quotidiennement au sein des différents projets de l’association sur des volets de communication, de recherche de fonds et plus particulièrement autour du volet scientifique.
Tu as été l’instigateur et le directeur scientifique de la mission scientifique de cette année dans le massif du Makay. Peux-tu nous raconter comment est né ce projet ?
Dessin d’Alix Thiebault de Gaëtan et d’un membre de l’équipe réalisant des échantillonnages (Source : G. Deltour)
Ce projet a démarré par ma volonté de mettre en pratique ce qui m’avait été enseigné durant mon parcours scolaire et ma formation au MNHN. Pour cela, j’ai formé une petite équipe au sein de mon Master et nous avons proposé de tester un outil interactif d’identification des Lémuriens durant une mission dans le Makay : Clémurs.
Petit à petit, mon équipe a été rejointe par d’autres étudiants et d’autres chercheurs qui souhaitaient apporter leurs compétences à la mission. Et celle-ci est devenue une mission scientifique pluridisciplinaire et intergénérationnelle. Mon rôle a alors été de travailler en collaboration avec l’ensemble des scientifiques et de mettre en place des protocoles de terrain.
Quelles ont été les difficultés rencontrées dans l’organisation : techniques, matérielles et humaines ?
Les difficultés sont liées à l’inaccessibilité du site. Il a fallu de nombreuses heures de bus, de 4×4 et de marche pour atteindre le massif, ce qui a beaucoup joué sur le moral de tous ! Le Makay est un massif qui se mérite ! Un des principaux problèmes a été de gérer l’acheminement du matériel scientifique en raison d’un manque de porteurs. En effet, la priorité était donnée à la nourriture et aux affaires personnelles des participants. Il a fallu attendre de longs jours avant que l’ensemble du matériel puisse être accessible à l’ensemble de l’équipe.
Ton retour dans le monde « moderne » remonte à quelques semaines, quel bilan fais-tu de ces six semaines d’expédition ?
Mon retour dans le monde « moderne » me fait à nouveau prendre conscience du rythme dans lequel nous vivons. Le décalage avec Madagascar est toujours intense. Après quelques carences en vitamines et en protéines, les repas à l’occidentale font beaucoup de bien !
Pour faire un bilan de cette expédition, ce sont près de 40 espèces animales et végétales qui viennent d’être recensées pour la première fois dans le Makay (lire encadré ci-dessus). Cependant, les découvertes auraient pu être beaucoup plus nombreuses sans les couacs logistiques. Il en ressort de mon point de vue que c’était une belle aventure !
Des prochains projets dans le Makay sont déjà en discussion. Est-il possible d’en savoir davantage ?
En effet, nous souhaitons poursuivre nos actions dans le massif du Makay autour de différentes thématiques.
Tout d’abord, au niveau scientifique, un projet de station permanente est en cours de réflexion. Un des problèmes du massif est son inaccessibilité. Il est nécessaire de proposer une structure pour les chercheurs qui souhaiteraient poursuivre des travaux dans ce site. Les précédentes expéditions scientifiques ont soulevé le potentiel du lieu mais elles doivent désormais laisser place à des recherches sur le long terme.
Des projets autour de l’amélioration des conditions de vie des populations locales avec un programme d’apiculture et le développement de l’éducation sont en cours.
Le développement de l’éco-tourisme dans la région doit aussi se faire en faveur des populations locales en leur fournissant de nouvelles sources de revenus.
Pour finir, comme souvent dans Passion-entomologie, aurais-tu une anecdote à faire partager aux lecteurs ?
Feu de brousse à la sortie du massif du Makay (Source : G. Deltour)
J’aimerais en partager beaucoup plus !!! Durant la mission, j’ai eu la chance de fêter mes 23 ans au sommet du Makay. Nous avons passé la nuit en bivouac sur une des crêtes du massif et nous avons vu le soleil se coucher et se lever en passant par une magnifique nuit étoilée ! C’était juste incroyable ! Je partage une seconde anecdote car celle-ci m’a beaucoup touchée. Sur le trajet du retour, installé dans un 4×4, nous sommes passés à proximité d’un gigantesque feu de brousse. Nous étions tellement choqués, que l’équipe est restée près de 30mn à regarder arbres et buissons brûler à quelques mètres de nous.
Remerciements
Photo du groupe avant le départ du camp Mahasoa (Source : B. Gilles)
Je souhaite vivement remercier l’ensemble des personnes qui, en ayant contribué matériellement et financièrement, m’ont permis de participer à cette mission (liste des personnes ci-dessous).
Je remercie également tous mes camarades d’infortunes croisés durant mon séjour et avec qui j’ai passé de très bons moments !
Liste des personnes ayant contribues matériellement et financièrement :
Ma femme qui m’a laissé la possibilité de partir !
Des gènes à l’origine de traits nouveaux donnant accès à un nouvel environnement
chez la punaise Rhagovelia (Heteroptera, Gerromorpha, Veliidae)
Les innovations évolutives se traduisent par l’apparition de nouvelles structures ou comportements au cours de l’évolution (ex : la fleur, la carapace des tortues, l’aile des insectes (lien), un patron de coloration chez les papillons (lien)). La théorie prédit que ces événements d’apparition sont uniques à chaque innovation et rares. C’est ainsi que les innovations sont partagées par des groupes d’espèces apparentées, des taxons (ex : respectivement, les angiospermes, les oiseaux, les tortues, les insectes, les papillons). L’apparition de nouvelles structures est le premier mécanisme par lequel les innovations participent à la diversification du vivant.
Une innovation est considérée comme clé, importante, lorsqu’elle permet l’adaptation à une nouvelle niche écologique via une nouvelle fonction. La théorie prédit que l’opportunité écologique ainsi créée va permettre la diversification de l’espèce possédant l’innovation en groupe d’espèces. C’est le deuxième mécanisme par lequel les innovations participent à la diversification du vivant. Ainsi, elles façonnent les trajectoires évolutives des lignées dans lesquelles elles apparaissent.
L’importance de certaines de ces innovations pousse les chercheurs à se demander quels sont les mécanismes génétiques et développementaux à leur origine. Ils cherchent à faire le lien entre les changement génétiques, phénotypiques et adaptatifs à un nouveau milieu.
Punaises du genre Rhagovelia à la surface de l’eau (Source : A. Khila, 2017)
Pour tenter de répondre à ces questions, l’équipe de Khila Abderrahman (CNRS-UMR5242, Université de Lyon) a étudié les mécanismes génétiques et les pressions environnementales qui sous-tendent l’évolution de « l’éventail propulseur » (photo 2 ci-dessous), structure exclusive des punaises du genre Rhagovelia (Heteroptera, Gerromorphe, Veliidae) qui leur permet de se propulser à la surface de l’eau.
Leurs résultats mettent en évidence le rôle central de gènes qui sont spécifiques au taxon. L’un d’eux n’est présent que chez les punaises d’eau et l’autre que dans le groupe qui possède l’éventail (le genre Rhagovelia). Ces deux gènes sont impliqués dans le développement de l’éventail. De plus, l’utilisation de tests biomécaniques a prouvé le rôle essentiel de l’éventail dans l’adaptation à un nouvel environnement, agissant ainsi comme une innovation déterminante au plan évolutif.
Biologie et description des Rhagovelia
Le genre Rhagovelia (photo 1 ci-dessous) est constitué de ∼200 espèces dont le cycle biologique se déroule, en général, à la surface de petits courants d’eau rapides – niche écologique pas ou peu accessible à la plupart des autres insectes semi-aquatiques -. Ces punaises aquatiques se déplacent à la surface par des mouvements d’aviron de la paire de pattes médianes où se situent les éventails. Un éventail se compose d’environ 20 branches (ou fanes) primaires chacune, dotée de branches secondaires plus minces (photo 2 ci-contre). Contrairement à Rhagovelia, le genre Stridulivelia, qui lui est étroitement apparenté, ne possède pas d’éventail de propulsion. Bien que les espèces des deux genres occupent les mêmes cours d’eau (espèces sympatriques), les punaises du genre Stridulivelia restent statiques sur les feuilles des plantes et n’effectuant qu’occasionnellement des déplacements rapides (voir vidéo 1).
Photo 1 : Rhagovelia antilleana – les éventails sont visibles sur les pattes médianes (Source : Sciencesnews.org)
Photo 2 : éventail propulseur chez Rhagovelia antilleana (Source : Santos et al., 2017) (Modifié par B. gilles)
Le développement embryonnaire de l’éventail débute entre 144 et 210 heures, soit à 35% de la durée totale de l’embryogenèse (à 26°C). Les larves du premier stade éclosent avec des éventails entièrement fonctionnels, renouvelés au cours de chacune de leurs cinq mues, indiquant la persistance du programme de développement du éventail tout au long des stades post-embryonnaires (illustration 1 ci-dessous).
Illustration 1 : Développement de l’éventail au 1er, 3ème et 5ème stade larvaire – augmentation de la taille et du nombre des branches (Source : Santos et al., 2017)
Analyses génétiques
Pour identifier les gènes à l’origine du développement de l’éventail, l’équipe de scientifiques a réalisé un profilage transcriptomique (étude de l’ensemble des ARN messager produits lors du processus de transcription d’un génome) des pattes embryonnaires chez Rhagovelia antilleana. L’étude de l’expression de ces ARNm a permis d’identifier 5 gènes s’exprimant uniquement dans la paire de pattes médianes et à l’endroit où se développe l’éventail : y (yellow), cp19 (cuticular protéine 19), ccdc174 (coiled-coil domain-containing protéine 174) et deux gènes similaires inconnus (c67063_g1 et c68581_g1, respectivement nommé geisha et mother-of-geisha) (illustration 2 ci-dessous). Aucun de ces gènes ne s’exprime dans les pattes antérieures et postérieures de R. antilleana ou de toute autre espèce apparentée comme Stridulivelia tersa et Oiovelia cunucunumana.
Illustration 2 : Expression des 5 gènes à l’apex de la patte médiane de Rhagovelia (Source : Santos et al., 2017)
Une recherche d’homologie des séquences liées à geisha et mother-of-geishaa montré que mother-of-geisha possédait un paralogue (gène homologue) retrouvé chez 5 espèces d’Hémiptères (punaises) et une d’Isoptères (termites). Aucun de ces deux gènes ne possède d’homologue dans d’autres Ordres comme les Lépidoptères, les Diptères et les Hyménoptères. Le gène mother-of-geisha possède cependant de proches homologues dans les espèces de punaise d’eau n’ayant pas d’éventail (i.e. autres que Rhagovelia) alors que toutes les espèces de Rhagovelia et seulement celles-ci partagent le gène geisha. Les chercheurs concluent que mother-of-geisha a émergé chez une espèce ancestrale commune à tous les Hémiptères et s’est ensuite dupliqué chez l’ancêtre commun du clade des Rhagovelia pour donner geisha. Dans cet ancêtre commun, la séquence de geisha a évolué très rapidement et a divergé de celle de mother-of-geisha (illustration 3 ci-dessous). C’est ainsi que les gènes ont des séquences très proches mais facilement différentiables dans toutes les espèces de Rhagovelia.
Illustration 3 : reconstruction phylogénétique des deux gènes (Source : Santos et al., 2017) (Modifié par B. Gilles)
L’examen de l’expression de ces deux gènes a révélé que geishaétait exprimé trois fois plus fortement que mother-of-geishadans les pattes médianes. Ainsi, non seulement la séquence protéique de ces gènes a divergé, mais aussi leur niveau d’expression.
Sachant que mother-of-geisha et geisha sont respectivement spécifiques des Hémiptères et des Rhagovelia et qu’ils sont exprimés à l’endroit et au moment du développement de l’éventail, les scientifiques ont souhaité tester s’ils étaient impliqués dans le développement de cette structure. Pour cela, ils ont inhibé l’expression de ces gènes lors du développement des larves. Ainsi, l’inhibition de l’expression des gènes mother-of-geisha et geishaa entrainé de la réduction à la disparition de l’éventail, tout en permettant le développement d’individus viables sans autres modifications phénotypiques d’autres structures (y compris des griffes directement liées à l’éventail) (Illustration 4 ci-contre).
Illustration 4 : Comparaison des phénotype sauvage et suivi d’une inhibition des gènes (Source : Santos et al., 2017) (Modifié par B. gilles)
La suppression du gène y « yellow » a eu pour conséquence le développement du phénotype « éventail jaunâtre » au lieu de couleur brun foncé. Ce gène connu pour intervenir dans la formation de pigments foncés, et conservé au sein du genre Rhagovelia, a donc été recruté pour assombrir la cuticule des éventails.
La surpression des gènes cp19 et ccdc174 connus pour leur implication dans le développement de la cuticule n’a quant à elle pas engendré de perturbation détectable pour des raisons non identifiées.
En résumé, dans cette partie les chercheurs ont établi un lien entre des changements génétiques (apparition de mother-of-geisha, duplication de geisha, évolution rapide de la séquence de geisha)et la génétique du développement de l’éventail (ces deux gènes sont fortement exprimés dans le territoire présomptif de l’éventail, leur expression est nécessaire au développement de l’éventail, yellow a été recruté lors du processus évolutif). Pour finir de reconstruire l’histoire évolutive ils ont tenté de comprendre en quoi ces changements étaient adaptatifs.
Mots de Abdourahman Khali
A. Khila sur le terrain (Source : A. Khila)
Vous dirigez l’Unité de recherche « Génomique du développement et évolution », quelles sont les thématiques et les modèles d’études que vous étudiez ?
Je dirige ce laboratoire au sein de l’UMR5242 regroupant 12 autres laboratoires. Dans mon laboratoire, nous nous intéressons aux mécanismes génétiques, évolutifs et écologiques à l’origine de la diversification des espèces. Les modèles d’études sont multiples : le poisson zèbre (Danio rerio), la drosophile (Drosophila melanogaster), la souris domestique, le tribolium (Tribolium castaneum – Coleoptera) et les Gerromorphes (punaises aquatiques).
Quels sont les outils innovants que vos équipes utilisent dans le cadre de leurs recherches ?
Nos équipes utilisent notamment une plateforme de séquençage à haut débit (PSI), le génie génétique et la manipulation des génomes, l’imagerie et microscopie, la modélisation, etc.
Que vont apporter vos recherches fondamentales en entomologie ces prochaines années et décennies ?
L’apport principal est d’accroitre nos connaissances scientifiques. D’autres apports peuvent toucher la sensibilisation du public par rapport aux écosystèmes et leur sauvegarde, et à un degré moindre, la lutte contre les insectes nuisibles.
Etudes du déplacement et du comportement
Un des objectifs majeurs de la biologie évolutive est non seulement d’identifier les mécanismes génétiques sous-jacents aux caractères nouveaux, mais aussi de déterminer l’impact des innovations sur l’histoire évolutive de l’organisme. Afin de déterminer si l’innovation éventail a permis l’adaptation à une nouvelle niche, les chercheurs ont testé si elle pouvait avoir participé à l’invasion des petits cours d’eau où vivent les Rhagovelia. Ainsi, ils se sont amusés à comparer les performances de locomotion par vidéographie à haute vitesse des déplacements à la surface de l’eau d’individus de Rhagovelia : 1) normaux (avec éventail), 2) sans éventail (ablation chirurgicale), 3) avec éventail rudimentaire (inhibition des gènes mother-of-geisha et geisha), 4) Stridulivelia tersa, une espèce proche sans éventail, qui vit aussi sur près de cours d’eau mais s’aventurant rarement à la surface.
Dans un premier temps, les chercheurs ont simplement confirmé que seul l’éventail était affecté par l’inhibition des gènes mother-of-geisha et geisha, et que les rudiments restants comme les griffes se déployaient et se rétractaient normalement.
Les scientifiques ont réalisé des comparaisons des performances locomotrices à la surface d’eau d’individus de ces 4 groupes. La première comparaison a été faite en eau stagnante. Pour cela, Stridulivelia tersa y sont plus rapides que les 3 groupes de Rhagovelia qui sont tous les 3 au même niveau. Cependant, Stridulivelia tersa et les Rhagovelia sans éventail font plus de battements de pattes par seconde. Les chercheurs concluent que sur l’eau stagnante les éventails ne sont pas utilisés pour augmenter la vitesse moyenne mais permettent de diminuer la fréquence des battements.
Ensuite, une deuxième expérience a consisté à obliger les groupes d’individus à ramer contre un courant d’eau lent (0,2m/s) et rapide (0,3m/s), simulant l’environnement des R. antilleana. Bien que les individus de S. tersa aient été plus rapides en eau stagnante, ils ont été emportés par les courant lents et rapides. Les individus R. antilleana normaux (témoins) ont quant à eux remonté les deux types de courant sans difficulté. De manière cohérente, les individus R. antilleana, ayant subi l’ablation de leurs éventails, n’ont pas réussi à remonter le courant rapide, et leur performance a fortement diminué dans le courant lent. Les individus aux éventails rudimentaires ont eu une performance intermédiaire. En fait, l’analyse de la fréquence de battement montre que les animaux ayant subi une ablation de l’éventail ou à l’éventail rudimentaire tentent de compenser en battant plus de fois par seconde. Dans certain cas cela suffit, mais pas toujours.
Les chercheurs concluent ainsi que l’éventail permet d’augmenter l’efficience des battements en augmentant la surface de la patte en contact avec l’eau et l’efficacité de chaque poussée par le transfert d’une plus grande quantité d’énergie. Ainsi, moins de battements sont nécessaires en eau stagnante, et des vitesses de pointes plus rapides permettent de lutter contre le courant.
Les performances de locomotion n’ont pas été affectées par le sexe des individus.
Enfin, les scientifiques ont évalué la capacité à effectuer des virages serrés, un comportement caractéristiques des espèces de Rhagovelia. Ils ont constaté que les virages des individus normaux (témoins) étaient systématiquement initiés par le déploiement des éventails et se réalisaient avec un angle moyen de 77°. Les individus des deux groupes sans éventails (inhibition et ablation) ont quant à eux tourné de manière moins efficace avec un angle intermédiaire de 111° et 126° respectivement.
Pris ensemble, ces résultats montrent l’importance de l’éventail dans le mode de vie de Rhagovelia. Plus important encore, un éventail rudimentaire confère toujours un avantage comparé à son absence. Les chercheurs pensent ainsi u’un éventail rudimentaire est apparu à la base des Rhagovelia puis il a été graduellement amélioré par une concordance entre des changements génétiques et la sélection.
Conclusion
Les résultats montrent le lien entre des mécanismes génétiques du développement, évolutifs et écologiques lors de l’émergence d’innovations comme l’éventail.
Punaises du genre Rhagovelia antilleana à la surface de l’eau (Source : A. Khila, 2017)
L’ordre phylogénétique de ces événements moléculaires demeurent inconnu. Les chercheurs ont reconstitué la majeure partie de l’histoire évolutive de l’éventail et proposent le scénario suivant. Initialement, le gain de fonction d’anciens programmes de développement dans la seconde paire de pattes a permis l’apparition d’un éventail rudimentaire. Puis, la duplication du gène geisha et son implication dans le développement a amélioré l’éventail. Chacune des étapes a pu être sélectionnée au cours de l’adaptation à la nouvelle niche que représentaient les cours d’eau. Cette étude apporte aussi un exemple, expérimentalement démontré et clair, de comment une nouveauté évolutive peu émerger graduellement à partir de structures plus simples ; une question clé en évolution.
Source
Santos M.E., Le Bouquin A., Crumière A.J.J. & Khila A. (2017) : Taxon-restricted genes at the origin of a novel trait allowing access to a new environment. Sciences, 358:386-390 (lien)
– The Insects: Structure and Function(R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
L’été, les rives du lac Mono en Californie sont envahies pas des millions de petites mouches. La densité est telle qu’elle atteint parfois par endroit plus de 2000 mouches sur la surface d’une carte postale, pour une population totale estimée à plus de 100 milliards d’individus. Ces mouches appartiennent à une espèce : Hephydra hians (Hephydridae). Les adultes sont connus pour leur mode de vie et leur comportement subaquatique des plus surprenants et singuliers : ils ont en effet la capacité à s’immerger complètement sous l’eau à la fois pour se nourrir et pour pondre leurs oeufs.
Les premières observations et descriptions des moeurs de cet insecte remontent à 150 ans et ont été faites par l’écrivain Mark Twain (1835-1910) sous ces mots : « vous pouvez les maintenir sous l’eau aussi longtemps que vous le souhaitez – sans que cela les dérange – elles sont seulement fières d’elles. Quand vous les laissez partir, elles remontent entièrement sèches à la surface, et marchant avec autant d’insouciance que si elles avaient été éduquées dans le but d’offrir un divertissement instructif à l’homme« .
Biologie et écologie de Hephydra hians
Hephydra hians est une de 4 à 7mm, de couleur marron foncé avec des reflets verts métalliques sur le thorax. L’espèce à la particularité de se développer dans des eaux à forte salinité, expliquant son nom de mouche alcaline. L’eau du lac Mono possède en effet une teneur en sel trois fois supérieure à celle de l’Océan Pacifique, avec une forte concentration en bicarbonate de sodium (NA2CO3), qui lui confère une alcalinité particulière (pH=10). Cette salinité résulte du non renouvellement de l’eau depuis 60 000 ans et d’une évaporation régulière et constante. Le calcium (CaCO3) qui l’accompagne provient quand à lui de sources naturelles.
Hormis H. hians, seuls des algues, des bactéries et des crevettes halophiles tolèrent cet environnement. Les mouches adultes, vivant entre 3 et 5 jours, peuvent ainsi s’alimenter d’algues et pondre sans craindre la concurrence ou la prédation. Le rôle écologique de ce lac est très important dans cette région car il offre une ressource alimentaire à près de 2 millions d’oiseaux migrateurs qui y font halte chaque année (par exemple, 85% de la population de goéland de Californie rejoignent le lac pour nicher) (voir vidéo en bas de page).
Hephydra hians dans sa bulle d’air (Source : F. van Breugel)
Il est primordial pour ces mouches de demeurer sèche pour leur survie. Le fait d’être mouillées engendrerait le dépôt d’une fine couche de minéraux sur leur cuticule (squelette externe des insectes), augmentant par conséquent la probabilité d’être à nouveau mouillées lors d’un prochain contact avec l’eau. Pour se protéger de l’élément liquide nocif lors de la plongée, les mouchent s’entourent d’une bulle d’air protectrice englobant à la fois le corps et les ailes. Cette boule leur permet ainsi de se protéger des sels et des composants alcalins et d’apporter l’oxygène tel un poumon externe (photo ci-contre). Grâce à ce scaphandre, la mouche peut rester immergée près de 15mn à des profondeurs de 4 à 8m. Pour rejoindre la surface, elle lâche prise et se laisse tout simplement porter où elle va pouvoir flotter, marcher sur l’eau ou encore s’envoler (voir vidéo ci-dessous).
Or, depuis 150 ans, la formation de cette bulle d’air et la capacité de Hephydra hians à s’immerger dans ces eaux aussi alcalines sont demeurées un mystère pour la science. C’est pourquoi, deux scientifiques du département de Biologie de l’Institut Technique de Californie (ITC), Floris van Breugel et Michael Dickinson ont mené des études les propriétés physico-chimiques uniques de la cuticule de ces mouches à l’origine de la formation de la bulle d’air (en partie financées par le National Geographic Society).
Adaptation à la plongée
La cuticule des mouches de la famille des Hephydridae est recouverte par de nombreux poils minuscules (setae) et par des cires (hydrocarbures).
Hephydra hians dans sa bulle d’air (Source : F. van Breugel)
Pour mieux comprendre les phénomènes physico-chimiques à l’origine du pouvoir hydrophobique des poils, les scientifiques ont d’abord observé au microscope l’aspect et la répartition des poils à la surface de la cuticule. Ils ont constaté que ces mouches possédaient un tapis de poils plus denses de 36% sur leur corps et leurs pattes que d’autres espèces de mouches apparentés (34% sur les ailes, 44% sur le thorax, 47% sur l’abdomen) (voir photo ci-contre).
Puis, pour mesurer les forces de tension à la surface de l’eau et de la cuticule de ces insectes, ils ont construit un capteur minuscule et plongé des centaines d’individus de H. hians, et de 6 autres espèces apparentées, dans une série de solution salines différentes en en faisant varier la salinité, le pH et la densité. La formation de la bulle d’air à la surface de la cuticule a pu être observée à l’aide de caméras à haute fréquence et de logiciels d’analyses mathématiques.
Michael Dickinson sur le terrain (Source : F. van Breugel)
Les résultats montrent que seule H. hians possède la capacité à former et à maintenir une bulle d’air dans des eaux aussi riches en bicarbonates de sodium (NA2CO3). Cette capacité ne résulterait que de quelques changements mineurs dans les propriétés physico-chimiques des hydrocarbures et des poils. Les propriétés détachante et de nettoyage de NA2CO3 sont connus de longue date (solution utilisée dans l’Egypte antique pour embaumer les momies). Comme le signal Mickael Dickinson, l’eau du lac est plus adaptée au lavage du linge que pour y vivre ! Cette adaptation a probablement évoluée au cours du temps avec l’augmentation lente et progressive de concentration en sels minéraux comme NA2CO3 dans le lac Mono (et dans d’autres lacs de la planète).
Récemment, les pressions de sélection sur les mouches alcalines du lac Mono sont devenues plus fortes. En raison d’une politique de détournement des eaux de la Sierra de l’Est menée par la ville de Los Angeles entre 1941 et 1982, la concentration en sels du lac a doublé. Cependant, les scientifiques ont démontré que cette augmentation assez soudaine en ions affectait plus les larves que les adultes.
L’adaptation principale ayant permis à H. hians d’accéder à cet environnement alcalin ne repose pas tant sur les propriétés physico-chimiques de ses poils ou de sa cuticule que sur l’apparition des comportements de déplacement et d’alimentation subaquatiques. Les scientifiques suggèrent que ces comportements ont pu apparaître lors de périodes où la disponibilité en nourriture en surface est devenue limitante, les algues sous-marines offrant une alternative abondante. Par l’absence de compétition et de prédateurs comme les poissons, la pression de sélection sur ces comportements subaquatiques est restée quant à elle plutôt faible. La bulle d’air ne couvre pas les yeux rouges de la mouche pour que sa vision ne soit pas déformée (photo ci-dessus), et ses pattes sont munies de griffes plus longues que celles d’espèces apparentées afin de lui permettre de se déplacer sur le fond au voisinage des berges du lac (voir vidéo ci-dessous).
Menaces
L’environnement de lac Mono est fragile et en danger par l’apparition de deux phénomènes : 1) Dérivation des eaux en amont du lac qui augmente la concentration en sel à des niveaux au-delà du seuil de tolérance de la mouche Hephydra hians ; 2) Formation d’un film d’écran solaire sur les eaux provenant des nageurs occasionnels qui altère les propriétés physico-chimiques des poils cireux.
Vidéos complémentaires
Source
van Breugel F. & Dickinson M.H. (2017) : Superhydrophobic diving flies (Ephydra hians) and the hypersaline waters of Mono Lake. Procceding of the National Academy of science, 1-6 (lien)
L’univers des insectes, en plus de fournir des sujets d’étude à l’entomologiste, constitue un véritable puit de ressources dans des domaines divers : traitement biologique des matières organiques, source de protéines pour l’alimentation animale, et épicerie d’ingrédients pour la cuisine humaine. Depuis quelques années, ce dernier domaine est de plus en plus sous les feux des projecteurs…
Insectes dans un marché en Thaïlande (Source : Google)
Cet intérêt a sa logique : d’un point de vue zootechnique et environnementale, l’élevage d’insectes (entomoculture) pour l’alimentation humaine a de quoi séduire. En effet, d’après le rapport de la FAO (Food and Agriculture Organization) de 2013, il nécessitera beaucoup moins d’intrants que les autres animaux : pour produite 1kg de grillons, on aurait besoin de 1,7kg de nourriture, contre 2,5kg pour les volailles, 5kg pour les porcs et 10kg pour les vaches (1). De plus, d’après ce même rapport, les vers de farine (Tenebrio molitor), les criquets et les grillons émettraient 10 fois moins de gaz à effet de serre et de nitrates par kilogramme de biomasse produite que le porc, et 25 fois moins que la vache. Enfin, ces espèces d’insectes vivent naturellement en populations à forte densité : il serait donc possible d’en élever de très grandes quantités hors sol sur une petite surface – pourquoi pas en milieu urbain – tout en leur assurant des conditions de vie respecterant leur bien-être.
D’un point de vue nutritionnel, consommer des insectes serait également cohérent : des analyses portant sur six espèces : grillon (Acheta domestica), couvain d’abeille, ver de farine (Tenebrio molitor), chenille mopane (Gonimbrasia belina), ver du palmier (Rhynchophorus ferrugineus) et ver à soie (Bombyx mori), ont montré qu’elles constitueraient des substituts intéressants à la viande. En effet, même si les valeurs nutritionnelles varient beaucoup d’une espèce à une autre, le modèle Ofcom, utilisé en Grande-Bretagne pour noter le caractère sain des aliments, a jugé ces espèces équivalentes au boeuf et au poulet d’un point de vue santé. Le système NVS (Nutrition Value Score) attribue même à ces insectes une meilleure note que la viande dans des contextes de sous-nutrition (2).
Vers à soie (Bombyx mori) en brochettes (Source : Google)
Aujourd’hui, la consommation d’insectes par les humains (entomophagie), fait surtout parler d’elle dans les médias dans deux contextes : comme divertissement (on pense à des émissions de télévision où manger des insectes est une épreuve voire une punition) ou comme « aliment du futur », le plus neutre possible, pour la valeur nutritionnelle de certaines espèces.
Les grands absents de cette représentation médiatique sont les consommateurs et consommatrices d’insectes dans des cultures autres que occidentales, qui les considèrent quant à eux comme ressources alimentaires, les récoltent, les élèvent et les cuisinent.
Acteur dans la production française d’insectes pour l’alimentation animale, Sébastien se demandait depuis longtemps comment les insectes étaient consommés autour du monde, quels modes d’élevage, quelles méthodes de collecte et quelles recettes de cuisine existaient déjà. Si le futur promet bien d’amener des insectes dans nos assiettes, autant apprendre à les préparer avec des spécialistes ! Il prépare avec sa compagne Annie un tour du monde des pays intégrant la consommation culturelle des insectes pour y réaliser des reportages et y chercher des recettes. Rentrés du Japon en novembre, ils repartiront pour le Cambodge en avril.
Sources :
Edible Insects – Future prospects for foods and feed security, Van Huis et al., 2013 – FAO (lien)
Are edible insects more or less « healthy » than commonly consumed meat ? A comparison using two nutrient profiling models developed to combat over- and undernutrition, Payne et al., 2016 – European Journal of Clinical Nutrition (lien)
Sébastien Collin
Annie Ruelle-Sanguine
Interview de Sébastien Collin et Annie Ruelle-Sanguine
Sébastien, peux-tu te présenter en quelques lignes ?
Nous sommes deux à travailler sur ce projet : Sébastien Collin et Annie Ruelle-Sanguine.
Je (Sébastien) suis ingénieur agronome en élevage animal, je me suis rapidement spécialisé dans l’élevage des insectes pour l’alimentation humaine et animale : au cours de ma formation et de ma vie professionnelle, j’ai pu travailler pour Jimini’s, EntomoFarm, Ynsect et le Département d’entomologie de l’Université de Sydney sur différents projets traitant de l’élevage de criquets et de vers de farine. Je suis aussi passionné de création audiovisuelle. Je réalise des vidéos sur internet depuis 2011 et des pièces radiophoniques depuis 2007.
Je (Annie) suis chimiste, ai fait un peu de prévention au travail, ai occupé trois postes d’enseignements en maths et en physique-chimie. Je fais actuellement de la médiation scientifique avec l’association « Les Petits Débrouillards ». J’écris aussi de la fiction et des articles depuis 2009.
Tu portes un projet sur l’entomoculture et l’entomophagie au niveau mondial, en quoi consiste-il ?
Notre projet, « Les Criquets Migrateurs« , a pour but de faire un tour du monde des pays où les insectes sont consommés, afin d’en apprendre plus sur les recettes traditionnelles, sur la façon dont ceux-ci sont collectés ou élevés, et sur la place qu’ils occupent dans les sociétés. Notre objectif est de réaliser des vidéos culinaires pour présenter ces recettes, ainsi que des documentaires pour les contextualiser dans leur culture d’origine. Nous allons aussi écrire un certain nombre d’articles sur notre blog afin d’approfondir ces sujets et tenir notre public informé de l’avancée du projet.
Grillons grillés au citron
Une petite recette à faire chez vous
Grillons grillés au citron
Faire revenir les insectes à la poêle avec de l’huile d’olive, quelques épices et du jus de citron, ce qui donne un amuse-bouche parfait à déguster au soleil.
20g de grillons
1/2 citron vert
Filet d’huile d’olive
Sel et poivre
Préparation :
Bien rincer les grillons avant utilisation
Ajouter l’huile d’olive, le jus de citron, le sel et le poivre aux grillons – bien mélanger
Verser la préparation dans une poêle – griller jusqu’à ce que ça croustille
Servir à l’apéro
Attention : ne pas consommer en cas d’allergies aux fruits de mer
Ce qui nous intéresse dans l’élevage d’insectes et son application en alimentation humaine, c’est l’impact culturel et gastronomique de ces ingrédients. Nous n’avons aucune culture de la consommation d’insectes en France pour l’instant : chez nous, tous demeure encore à inventer. Or, les médias nous parlent beaucoup des qualités nutritionnelles et environnementales des insectes, mais il reste encore un blocage mental chez beaucoup de gens à l’idée d’en manger. Nous pensons que le problème vient du fait que le public a du mal à voir les insectes comme des ingrédients qu’il pourrait utiliser au quotidien, chez lui, dans sa cuisine ; la santé et l’écologie sont des sujets importants, mais quand il s’agit de nourriture, ce sont peut-être des arguments un peu faibles pour se projeter dans une gastronomie basée sur l’entomophagie. C’est « l’aliment du futur » : une idée d’un mode d’alimentation amélioré, respectueux de l’environnement et bon pour notre santé, mais qu’on visualise plus dans un contexte de science-fiction et qu’on peine à imaginer dans l’immédiat.
Or l’une des principales raisons pour laquelle les insectes sont autant consommés dans le monde, c’est simplement qu’ils sont très bons. Deux milliards d’êtres humains voient les insectes d’abord comme des ingrédients délicieux avant d’être une source facile de protéines. Et c’est ce pont de vue que nous souhaitons faire partager !
Pour cela, nous allons adopter le format de la vidéo culinaire : rapide, dynamique, intéressante à regarder, c’est un mode de présentation des plats qui met en valeur les ingrédients en déconstruisant la recette pas à pas d’une façon simple. Tous ces ingrédients savoureux filmés en gros plans en train de cuire donnent fin au public, l’impliquent et lui inspirent l’envie d’essayer. Et c’est précisément ce que nous voulons faire : donner faim aux gens avec des insectes. Après « les insectes ? Oui il parait que c’est sain et écologique », nous voulons qu’ils se disent « les insectes ? Oui, ça à l’air bon et j’aimerais bien essayer ». Nous avons déjà fait une vidéo pilote présentant ce concept : elle est visible en cliquant ici.
Tu souhaites découvrir plusieurs pays, coutumes et cultures autour de l’insecte, comment s’organise ton périple ?
Nous travaillons sur ce voyage depuis mai 2017, pour un départ prévu début avril 2018 (nous avons déjà fait une petite expédition au Japon en novembre). Les insectes sont consommés dans plus de 110 pays : il nous aurait été difficile de tous les visiter. Nous en avons donc sélectionné huit, en répartissant notre itinéraire en fonction des saisons d’activités des espèces que nous voulons étudier :
Japon – début novembre 2017 (2 semaines) : visite du festival de la guêpe de Kushihara. Au Japon, les insectes les plus populaires sont le criquet, la guêpe, le frelon, le ver à soie et les zazamuchis (petites larves aquatiques – Trichoptères). Un article et une vidéo ont été publiés sur ce sujet ! Tout est visible sur notre site internet : lescriquetsmigrateurs
Nymphes et larves de guêpes poêlées – Japon (Source : S. Collin)
Cambodge – début avril à mi-mai 2018 (6 semaines) : nous espérons goûter aux fourmis tisseuses, aux criquets et surtout aux mygales frites. Ces dernières ne sont peut être pas des insectes, mais elles comptent quand même : les photos nous donnent beaucoup trop envie pour ne pas les inclure dans notre projet
Thaïlande – mi-mai à fin juin 2018 (6 semaines) : capitale mondiale de la consommation d’insectes. On y goûte une diversité étonnante, comme les criquets, les grillons, les vers de farine, les vers de palme (Rhynchophorus ferrugineus), les chenilles de bambou (Omphisa fuscidentalis), les vers à soie (Bombyx mori), les punaises d’eau… On aura de quoi faire !
Australie – juillet 2018 (un mois) : le mois de juillet est une excellent période pour trouver des « witchetty grubs », des chenilles de l’espèce Endoxyla leucomochla (famille des Cossidae) consommées occasionnellement par les Aborigènes et vivant dans les racines de l’arbre witchetty. Nous avons aussi lu que les termites étaient également consommés, donc nous espérons pouvoir en déguster aussi
Mexique – août 2018 (un mois) : dans l’état d’Oaxaca, dans le sud du pays, on récolte des criquets appelés localement chapulines et des larves de fourmis du genre Liometopum appelées escamoles. On peut les préparer en tacos ou avec du guatamole
France – septembre à mi-octobre 2018 (6 semaines) : en attendant la saison des pluies du Kenya, nous effectuerons une escale en France. Ce sera l’occasion de faire le point sur ce qui se fait en termes d’élevage d’insectes chez nous, en Europe : où en sommes-nous dans l’intégration de l’insecte dans les circuits agro-alimentaires ? Quels usages en fait-on, et où pouvons-nous en goûter ?
Kenya – mi-octobre à mi-novembre 2018 (4 semaines) : nous arriverons en début de la saison de récolte des nsenene, des sauterelles du genre Ruspolia très appréciées dans l’est de l’Afrique. Nous espérons aussi goûter aussi aux vers de palme et aux termites, toujours très populaires, ainsi qu’au kungu cake, un gâteau du lac Victoria fabriqué à partir de moucherons compressés
Botswana – mi-novembre à mi-décembre (4 semaines) : le Botswana sera un excellent endroit où découvrir la chenille de l’espèce Gonimbrasia belina, appelée localement mopane, un insecte populaire dans le sud de l’Afrique. Nous arriverons en début de saison, ce qui sera l’occasion d’en apprendre davantage sur sa récolte.
Planning du tour de monde de Sébastien Collin et de Annie Ruelle-Sanguine
Quels sont les moyens matériels et financiers nécessaires pour mener à bien ton projet ?
Nous allons avoir besoin d’un budget total de 28 000 €. Cela inclut les billets d’avion, les logements, la nourriture, les déplacements sur place… Nous avons aussi un peu de matériels : un ordinateur pour le montage vidéo, un micro USB et un caméscope qui tient plutôt bien la route.
Tu souhaites faire appel à la communauté Passion-Entomologie pour finaliser ton voyage, que te manque-t-il ?
Vous souhaitez aider et contribuer au financement et au prêt de matériels : n’hésitez pas !!
Sébastien et Annie recherchent pour finaliser leur projet de tour du monde de l’entomoculture
Ce projet sera majoritairement autofinancé, mais nous sommes à la recherche de sponsors afin de compléter notre budget. Nous sommes date soutenus par Jimini’s et EntomoFarm, qui nous financent à hauteur de 5 000 €. Si on ajoute nos propres fonds, nous avons de quoi aller au Kenya : il nous manque entre 2 000 et 3 000 € pour effectuer le trajet complet. Donc, si parmi les lecteurs et lectrices de Passion-Entomologie se trouve un.e dirigeant.e d’entreprise qui souhaiterait nous sponsoriser, nous sommes ouvert à la discussion ! Notre dossier complet peut être téléchargé à cette adresse.
Alternativement, nous serions aussi preneurs de matériel, si quelqu’un peut nous en prêter ! Nous pensons par exemple à une meilleure caméra, avec un micro performant. Et si vous avez des contacts dans les pays que nous allons visiter – de collecteurs/éleveurs/chercheurs/cuisiniers d’insectes – une mise en relation nous aiderait énormément.
Et, bien entendu, l’un des meilleurs soutiens que vous pouvez apporter au projet, c’est de le suivre, de nous dire ce que vous en pensez et de le partager autour de vous. Donc n’hésitez pas à aller faire un tour sur notre chaîne Youtube, sur notre site, et sur votre réseau social préféré (Facebook, Twitter, Instagram et même LinkedIn).
Y a t-il une chose qui te tienne à coeur et que tu souhaites absolument réaliser ?
Outre l’enrichissement gastronomique, il y a une application potentielle de la consommation d’insectes qui me fascinerait au plus haut point : celle de l’alimentation des astronautes. Développer des bases lunaires ou martiennes pourrait devenir une réalité plus rapidement que l’on ne le pense : il faudra alors trouver des modèles agricoles pertinents pour nourrir les habitants de ces bases. Il s’agit de développer des moyens de production qui ne prennent pas trop de place, qui ne consomment pas trop d’intrants, qui soient durables, qui fonctionnent en circuit fermé et qui permettent de nourrir les gens sainement : les insectes pourraient satisfaire ces exigences. Au passage, ce genre de modèle nous serait bien utile sur Terre ! La recherche spatiale permet souvent de développer des technologies qui trouvent une application en dehors de son domaine propre ; et on retrouverait cette tendance ici aussi : le moteur de l’innovation que représente cette recherche nous permettrait de répondre à des enjeux et les problématiques que nous rencontrons également sur Terre – et si les insectes peuvent faire partie de la solution, tant mieux !
Ouvrages et produits recommandés sur cette thématique :
Micromalthus debilis, un Archostemate (plus petit sous-ordre des coléoptères – 50 espèces) d’origine nord-américaine ne semble pas avoir évolué depuis plusieurs dizaines de millions d’années : il est l’unique représentant vivant connu de la famille des Micromalthidés.
Larves et adulte de M. debilis (Source : Bugguide.net)
Bien que l’espèce affiche des caractères morphologiques archaïques, son cycle de développement est quant à lui à la fois complexe et singulier comme a pu le révéler l’entomologiste américain Herbert S. Barber (1882-1950) – que ses observations en la matière mirent en doute sa crédibilité -. Depuis, seulement cinq expérimentations ont été réalisées sur cette espèce en cent ans.
Le cycle de M. debilis combine à la fois de l’haplodiploïdie, de l’uni-oviparité (un oeuf mâle pondu à la fois), de la poly-viviparité (plusieurs larves femelles sont délivrées à chaque fois) et de la matriphagie interne et externe (consommation de la mère par la descendance).
Tout d’abord, les populations de M. debilis sont exclusivement constituées de larves femelles (néoténie*) qui, chose surprenante, ne donnent naissance à leur tour qu’à des larves également femelles sans passer par le stade adulte. Ce mode de reproduction est appelé parthénogenèse thélytoque (lien) ou en encore pédogenèse. Il existe un dimorphisme entre les stades larvaires : au premier stade elles sont actives et très mobiles (triangulins – 1mm) ; et apodes au second (cérambycoïdes – 3,3mm).
* Néoténie : absence de la phase adulte, le cas le plus connu concerne une sorte de salamandre mexicaine, l’axolotl (Ambystoma mexicana). Les individus adultes possèdent des organes larvaires comme des branchies externes par exemple.
Larve au premier stade – Triongulins – de M. debilis (Source : Tolweb.org)
Larves au second stade – cérambycoïdes – de M. debilis (Source : Buggnit.net)
Encore plus surprenant, lorsque les conditions sont défavorables (si le bois dans lequel les larves se développent sèche trop par exemple), la larve au second stade a deux options : 1) Se nymphoses et donner un adulte ailé femelle, 2) Muer une nouvelle fois et livrer une larve de troisième stade également pédogénétique. Cette larve acquière alors la capacité de pondre par parthénogenèse un oeuf unique (arrhénotoque), mâle, puis sombre dans une véritable léthargie. De cet oeuf éclos une larve d’un autre type (curculionide), mâle bien sûr, très différente de la larve femelle : aussitôt, elle enfonce sa tête dans l’orifice génital de sa mère endormie pour… s’en nourrir (matriphagie). Le festin dure une semaine avant que la larve ne se nymphose et ne livre un mâle ailé. Il est possible que cette larve conduite à la matriphagie car elle n’est pas dotée des bactéries symbiotes permettant de digérer le bois.
Les adultes sont très rarement observés, que ce soit dans l’environnement ou en laboratoire. Des études récentes, menées par l’Anglais M. Alejandra Perotti et son équipe, ont mis en évidence que les adultes pouvaient apparaître, in vitro, lorsque des larves cérambycoïdes étaient soumises à des chocs thermiques. Ces chercheurs suggèrent que cet insecte ait quasiment perdu la capacité à se métamorphoser suite à des mutations au niveau de gènes régulateurs clés régulant cette transformation. Le passage à l’état adulte est particulièrement rare car les scientifiques n’ont pu obtenir qu’une seule femelle pour 650 larves et un mâle pour 10 000 larves cérambycoïdes !
Le fait que les mâles soient devenus anecdotiques dans les populations pout s’expliquer de trois manières : 1) Une diminution de la probabilité d’accouplements par un rejets des femelles pour les mâles apparentés et une faible capacité de dispersion de ces derniers, 2) Une compétition pour les ressources locales, 3) La progéniture d’un sexe augmente la fécondité et la valeur reproductrice (fitness) d’un des parents, ici, les mâles tuent et se développent au détriment des femelles, renforçant ainsi le biais du sex-ratio en faveur des femelles. Cependant, les dernières études ne permettent pas de démontrer et de confirmer l’une de ces théories.
Larves au second stade -cerambycoïdes – de M. debilis (Source : Tolweb.org)
D’après l’équipe de Perotti, le biais en faveur des femelles pourrait également s’expliquer par la présence d’un cycle haplodiploïdique. En effet, les populations de nombreuses espèces haplodiploïdes, sociales ou non, comme les guêpes parasitoïdes (lien), possèdent un sex-ratio biaisais en faveur des femelles. Des espèces parthénogénétiques haplodiploïdiques comme le sont Micromalthus debilis, les guêpes Cinipidae et les mouches Cecidomyidae, présentent des cycles similaires avec un biais en faveur des femelles, bien qu’elles soient non apparentées. Cependant, peu d’études se sont intéressées aujourd’hui à ces cycles biologiques.
L’étude de Perotti a confirmé expérimentalement que les adultes sont physiologiquement dans l’incapacité de se reproduire, cette perte étant probablement survenue au cours du temps par dégénération des rares adultes. Bien que des cas de mâles stériles chez d’autres espèces aient pu être décrits, le fait que chez Micromalthus debilis des femelles le soient également reste sans précédent d’après P.D.N. Hebert (1987). Le « coût » biologique que représente la « fabrication » de ces individus biologiquement inutiles expliquerait que l’essentiel du cycle ait évolué vers la parthénogenèse. Les adultes « fantômes » seraient ainsi des reliquats d’un cycle biologique ancestral sexué devenu aujourd’hui simplement vestigial.
Le processus à l’origine de la féminisation des populations peut également faire intervenir des bactéries endosymbiotiques (Rickettsia bellii), détectées chez toutes les larves analysées, connues pour modifier le sex-ratio de la population et induire une « féminisation » du cycle. Si ces bactéries rétablissent la diploïde et donc la formation de femelles, la chaleur permettrait de neutraliser ou de tuer les Rickettsia induisant la formation de mâles haploïdes. Aucun symbiote n’a en effet été détecté chez les adultes.
Quoiqu’il en soit, fort de cette biologie singulière, ce coléoptère « fossile-vivant » minuscule colonise depuis des temps immémoriaux les milieux les plus étonnants, comme les étais des galeries de mines ou les traverses de voie ferrée, en association avec des moisissures. Il a été ainsi introduit dans quasiment toutes les régions du globe.
Cycle biologique de Micromalthus debilis (Source : Perotti et al., 2016)
Source :
Perotti M.A. ; Young D.K. & Braig H.R. (2016) : The gost sex-life of the paedogenetic beetle Micromalthus debilis. Nature-Scientific Reports, 6:27364 (lien)
Voila plus de 160 ans que la tête orange d’un spécimen de la mouche Thyreophora cynophila n’avait pas été observée ! Cette espèce de la famille des Piophilidae (photos ci-dessous), décrite en 1798 par le botaniste et entomologiste G.W.F. Panzer (1755-1829), avait été observée pour la dernière fois près de Paris en 1840. Sa présence avait été constatée auparavant en Allemagne, en Autriche et en France.
Son absence durant toutes ces décennies lui a valu son inscription dans la liste des animaux européens considérés éteints, faisant de cette mouche l’unique représentant de l’ordre des Diptères tenu comme disparu et le seul cas constaté d’éradication par l’homme.
Sa réapparition constitue donc un événement exceptionnel. C’est durant une campagne d’études et de piégeages d’insectes sarcosaprophages, menée par l’entomologiste espagnol D. Martin-Vega dans la région de Madrid, qu’un jour de janvier 2007, lors de relevés de pièges, 6 individus de T. cynophila sont apparus aux yeux des chercheurs parmi des centaines de spécimens d’autres espèces de mouches.
Ces nouveaux spécimens viennent s’ajouter aux 16 autres connus à ce jour et présents dans les collections d’Europe, ces 8 mâles, 7 femelles et un sexe non déterminé, étant répartis dans sept Museum d’Histoire Naturelle européens : 4 à Paris, 4 à Vienne, 2 à Halle (Saale), 2 à Stockholm, 2 à Berlin, 1 à Copenhagen et 1 à Londres.
Spécimen de Tyreophora cynophila – février 2007 : A) Spécimen de profil – B) Spécimen de dessus (Source : Martin-Vega et al., 2010)
Biologie et écologie de Thyreophora cynophila
En plus de son apparence singulière (voir photos ci-dessus), le cycle de vie et la biologie de T. cynophila sont également étonnants à plus d’un titre. Les mouches adultes sont attirées par les carcasses de gros mammifères au stade de putréfaction (avec une préférence pour les ongulés comme les chevaux et les ânes), où les larves se développent dans les os de grande taille. Contrairement à la très grande majorité des autres espèces de mouches, le cycle phénologique (cycle lié aux conditions climatiques) de T. cynophila se déroule en hiver. Les individus, nocturnes, sont actifs entre les mois de janvier et de mars. Plusieurs sources, comme l’entomologiste et le diptériste français Justin Pierre Marie Macquart (1776-1855) en 1835, rapportent que la tête possèderait une capacité de bioluminescence. Le faible nombre d’observations fait que la biologie, le cycle de développement et l’écologie de cette espèce demeurent toutefois en très grande partie inconnus.
La disparition de ces mouches proviendrait de la somme de multiples événements. D’après Menier (2003), l’extinction possible de T. cynophila résulterait de la disparition des grands prédateurs, comme le loups, en Europe de l’Ouest, capables de briser et d’écraser les os de grande taille dans lesquelles les larves se développent, rendant inaccessible la principale source de nourriture de T. cynophila. Pour Linder (1949), la phénologie particulière de l’espèce la rendrait invisible aux yeux des entomologistes qui limitent leurs observations et leurs collectes durant l’hiver, et notamment sur les carcasses.
S’ajoute à cela, l’augmentation des activités humaines entrainant pollution, fragmentation des milieux, introduction d’espèces invasives et changement climatique qui bouleversent les populations d’insectes, voire leur disparition, bien que cela reste difficile à constater. Par exemple, Dunn (2005) estime que près de 44 000 espèces d’insectes auraient disparu au cours des 600 dernières années, alors que seulement 40 extinctions sont avérées !
La découverte de plusieurs spécimens de Thyreophora cynophila pose la question de la viabilité de la population et des mesures à prendre pour sa conservation et son maintien. Il s’agirait de préserver un environnement offrant une quantité de cadavres suffisante, chose difficile depuis la crise de l’encéphalopathie spongiforme bovine en Europe.
Comme l’a indiqué le diptériste russe Sergey Paramonov (1894-1967) en 1954, il est probable que l’espèce ne soit pas si exceptionnelle que cela dans la nature, mais seulement que leur collecte soit rare. Qui irait, en effet, collecter des mouches, la nuit, en plein hiver sur de gros cadavres ?
Vidéo :
Source
– Martin-Vega D. ; Baz A. & Michelsen V. (2010) : Back from the dead : Thyreophora cynophila (Panzer, 1798) (Diptera : Piophilidae) « Globally extinct » fugitive in Spain. Systematic Entomology, 35:607-613 (lien)
Les yeux (la vision), dans leur diversité de forme et de structure, permettent à un organisme, quel qu’il soit (invertébrés et vertébrés), de collecter l’énergie lumineuse (photons), de la convertir en énergie électrique qui sera ensuite transmise au cerveau via des neurones où elle sera analysée et interprétée pour donner naissance à une représentation mentale de l’environnement à l’animal lui permettant de se déplacer, de repérer une proie ou un partenaire sexuel.
Cependant, l’oeil des arthropodes est différent de celui des vertébrés tant au niveau de son anatomie que de son mode de fonctionnement. Cette série de trois articles essaie d’apporter une synthèse générale sur la vision chez les insectes : 1) Anatomie et structure des yeux ; 2) Vision et perception du mouvement ; 3) Physiologie des récepteurs et mécanismes de régulation de la sensibilité à la lumière.
I. Transduction de l’énergie lumineuse en énergie électrique
La conversion d’un signal lumineux en énergie électrique puis en message nerveux implique un mécanisme appelé phototransduction. Ce processus fait intervenir des récepteurs protéiques photosensibles : les pigments visuels, appelés rhodopsines. Chez les insectes, ces protéines se rencontrent dans de petites structures spécialisées : les chromophores, elles même présentent dans le rhabdomère des ommatidies (lire cet article).
Oeil composé et ommatidies d’un insecte (Source : Ecole Polytechnique Fédérale de Lausanne)
Par exemple, chez la drosophile (Drosophila melanogaster), 500 à 2000 molécules de rhodopsines sont présentent dans chacune des 40 000 microvilli que contient un rhabdomère d’une ommatidie.
Lorsqu’un photon (particule lumineuse) est absorbé par une molécule de rhodopsine, une cascade complexe d’événements chimiques entraine une dépolarisation (changement de charge électrique) de la membrane d’une cellule nerveuse (neurone).
La probabilité pour qu’un photon rencontre une molécule de rhodopsine dépend de la longueur du rhabdomère : ainsi, avec une longueur de 80µm, le rhabdomère de la drosophile absorbe 26% de la lumière contre 100% chez la libellule. Chez d’autres espèces, nocturnes ou évoluant dans la pénombre, un tapetum* (surface réfléchissante) renvoie la lumière, tel un miroir, une deuxième fois vers le rhabdomère, augmentant ainsi la probabilité de rencontre d’un photon avec une molécule de rhodopsine et donc la sensibilité de l’organe visuel à la lumière.
*Tapetum : le même principe se retrouve chez les yeux de vertébrés nocturnes comme les chats, leurs yeux reflétant la lumière des phares d’une voiture par exemple.
II. Adaptation aux changements de luminosité
Les cycles nycthéméraux entrainent de grandes fluctuations importantes de luminosité entre le jour et la nuit : 4×10.20 photons par mètre carré et par seconde en journée contre 10.10 photons durant la nuit (variation d’un facteur 40 milliards !). Afin de pallier cette contrainte environnementale, les insectes ont développé de multiples adaptations situées à deux niveaux : 1) Régulation de la quantité de lumière atteignant les photorécepteurs ; 2) Modification de la sensibilité des photorécepteurs.
II.1. Régulation de la quantité de lumière atteignant les photorécepteurs
La quantité de lumière qui atteint le rhabdomère est régulée par le mouvement des pigments dans les cellules, mouvement parfois associé à la modification anatomique des ommatidies. Cette adaptation se retrouve en majorité dans des yeux composés de type « superposition » (lire cet article) (Illustration 1 ci-dessous).
Illustration 1 – Formation de l’image : a) dans un oeil composé de type apposition, la lentille formant une image inversée de l’objet, b) dans un oeil composé de type superposition, les rayons lumineux sont réfractés à l’intérieur de la lentille (Source : The Insects : Structure and Function – R.F. Chapman – 5ème Edition (2013) – p716 – Modifié par B. Gilles)
Yeux de types superposition
Les adaptations se situent à deux niveaux :
Illustration 2 – Exemple d’adaptation à 1) la nuit (à gauche) – 2) à la lumière (à droite) (Chez le genre Archicauliodes, Mégaloptères) – D’après Walcott, 1975 (Source : The Insects : Structure and Function – R.F. Chapman – 5ème Edition (2013) – p711 – Modifié par B. Gilles)
– Pigments : dans un environnement obscur, les pigments sont situés dans la zone distale des cellules pigmentaires, permettant ainsi aux photons de se propager entre les ommatidies : zone claire (Illustration 2 ci-contre – cliquer dessus pour agrandir). A l’inverse, en présence d’une forte intensité lumineuse, les pigments se déplacent vers le centre des cellules afin de limiter l’entrée de lumière en diminuant la quantité de photons pénétrant dans l’ommatidie. A ce moment, le fonctionnement de l’oeil est similaire à celui de type « apposition », où chaque ommatidie fonctionne indépendamment des autres (lire cet article).
Un procédé similaire existe dans nos yeux, le cristallin se ferme ou s’ouvre plus ou moins en fonction de l’intensité lumineuse présente dans l’environnement.
– Cellules photoréceptrices : l’adaptation au changement de luminosité se réalise par l’extension et la compression du cône cristallin, cantonnant le pigment aux parties périphériques de l’ommatidie. En présence d’une forte luminosité, les cellules photoréceptrices sont courtes et les cellules pigmentaires primaires s’étendent sous la lentille, ces déplacements peuvent représenter près de 15µm.
Les adaptations à une variation de luminosité requièrent parfois plusieurs minutes, voire davantage. Chez les fourmis Camponotus, les premiers changements interviennent 15 minutes après un changement d’intensité lumineuse pour se finir 2 heures après. Il existe une grande variété de tempo entre les espèces. Ainsi, chez les papillons de nuit (hétérocères) du genre Cydia (des Carpocapses), le processus complet, d’une durée de 1 heure, débute 1h30 avant le crépuscule ou l’aurore pour que l’adaptation soit terminée 30 min avant le changement total de luminosité.
L’intensité lumineuse perçue est également dépendante de l’habitat dans lequel l’insecte évolue. Par exemple, le coléoptère cavernicole du genre Zopholas perçoit une luminosité de magnitude 5 fois plus faible que des espèces diurnes de la même famille (Tenebrionidae), comme le genre Tenebrio.
Yeux de type apposition
Illustration 3 – Changements structuraux dans une ommatidie d’un oeil de type apposition chez une fourmis Camponotus – seule la partie distale est visible (d’après Menzi, 1987) (Source : The Insects : Structure and Function – R.F. Chapman – 5ème Edition (2013) -p725 – Modifié par B. Gilles)
Des mouvements pigmentaires comparables aux yeux de type superposition sont observés dans ceux de type apposition.
Par exemple, chez la fourmi du genre Camponotus, les cellules pigmentaires primaires se contractent lorsque la luminosité augmente, comprimant le cône cristallin, formant un tractus cristallin étroit entouré de cellules pigmentaires primaires (Illustration 3 ci-contre). Ce rétrécissement diminue l’entrée de la lumière et seule la lumière arrivant dans l’axe de l’ommatidie atteint le rhabdomère. Chaque ommatidie est régulée de manière indépendante par rapport aux autres.
II.2. Modification de la sensibilité des photorécepteurs
La régulation de la sensibilisation des récepteurs s’opère, dans les yeux de type apposition, par un changement structurel des cellules photoréceptrices. Leur structure varie selon la disponibilité lumineuse absorbée par les molécules de rhodopsines.
Des granules de pigments absorbants sont présents à l’intérieur des cellules réceptrices entourant l’intérieur des microvilli du rhabdomère (Illustration 4 ci-dessous). Dans la clarté, les granules pigmentaires migrent vers la surface interne du rhabdomère et absorbent la lumière circulant le long de ce dernier, diminuant ainsi la quantité de lumière et l’intensité lumineuse reçues par les photorécepteurs. Dans l’obscurité, le processus s’inverse : les pigments absorbants migrent vers la périphérie.
Les mouvements de ces pigments, contrairement à ceux situés dans les cellules pigmentaires, sont quant à eux rapides : quelques secondes à peine suffisent par exemple chez les diptères (mouches) et les lépidoptères (papillons).
Illustration 4 – Changement dans les cellules photoréceptrices dans un oeil de type apposition – coupe transversale d’un rhabdomère (à gauche) – Coupe longitudinale avec trajectoire de la lumière (à droite) – D’après Snyder & Horridge, 1972) (Source : The Insects : Structure and Function – R.F. Chapman – 5ème Edition (2013) -p726 – Modifié par B. Gilles)
III. Sensibilité spectrale et vision chromatique
Les cellules photoréceptrices possèdent une gamme de sensibilité (profil Gaussien) à des longueurs d’onde comprises entre 330 et 640 nm (voir figure ci-contre). La longueur d’onde maximale dépend de la composition de la séquence en acides aminés de la protéine d’opsine.
Les ommatidies sont caractérisées selon la présence de certains types de récepteurs :
Type I : 1 récepteur aux UV et au bleu
Type II : 2 récepteurs aux UV
Type III : 2 récepteurs au bleu
L’étant ancestral des photorécepteurs chez les Ptérygotes (insectes ailés) semble être une sensibilité aux UV, au bleu et au vert. Ces types de récepteurs se retrouvent dans une grande proportion des ommatidies. Certaines espèces possèdent 6 récepteurs différents au vert. Ainsi, des hyménoptères (guêpes, abeilles et bourdons) et des lépidoptères des genres Vanessa ou Mandura sont qualifiés de tri chromatiques : capacité à percevoir 3 couleurs.
Chez les diptères, il existe deux autres types d’ommatidies : « pâle » et « jaune », composées de 6 récepteurs (R1 à R6) couvrant le spectre de 300 à 600 nm qui correspond,dent à une combinaison d’opsines et de pigments sensibles et absorbants les UV.
Chez de nombreux insectes, la proportion de différents pigments visuels varie selon la fonction que joue la vision. Par exemple, chez les abeilles, les ommatidies en position dorsale sont sensibles essentiellement aux UV et au bleu, alors que celles en position ventrale le sont au vert. Cette configuration se retrouve chez de nombreuses espèces volantes : l’oeil serait en fait constitué pour équilibrer l’insecte durant le vol par maintien de l’horizon au centre de la vision, ciel (bleu) et sol (vert). Il existe également des variations entre les sexes. Chez le papillon Lycaena rubidus, les femelles possèdent des ommatidies dorsales sensibles à des longueurs d’onde de 568 nm, absentes chez les mâles.
Plusieurs espèces de lépidoptères, d’odonates (libellules) et possiblement de quelques coléoptères et hyménoptères possèdent plus de 3 types de photorécepteurs sensibles à la couleur avec par exemple une sensibilité maximale pour le rouge ou pour le violet. Cette perception est rendue possible par une augmentation de la diversité de types d’opsines (duplication de gènes par exemple) ou par l’utilisation de filtres contrôlant la quantité de lumière atteignant les pigments, voire par une combinaison de ces deux stratégies.
La diversité spectrale de la réception visuelle peut être assurée au sein des cellules photoréceptrices de deux manières : 1) Posséder la même molécule d’opsine et 3 filtre pigmentaires différents entourant le rhabdomère, 2) Posséder 5 molécules d’opsines différentes et un filtre péri-rhabdomal, comme chez le papillon Papilio xuthus.
La sensibilité au rouge est obtenue par la présence d’une opine sensible à des longueurs d’onde proche de 575 nm, la même sensibilité que les récepteurs humains (565 nm). En règle générale, la sensibilité d’un récepteur à une couleur est directement liée à un type d’opsine.
Comme chez les diptères, les photorécepteurs de P. xuthus sont disposés en couches afin que la lumière atteignant les cellules photoréceptrices des couches inférieures soit filtrée par celles des couches supérieures. Chez d’autres insectes, la lentille est colorée de telle sorte que les rhabdomères reçoivent une lumière filtrée monochromatique (constituée d’une seule longueur d’onde). Par exemple, de nombreuses espèces de mouches possèdent à la surface de leurs yeux composés des lignes verticales ou horizontales de couleurs variables : mâle de la famille des Dolichopodidae, mouches de la famille des Tabanidae (taons) ou encore Hermetia illucens (Stratiomyidae) (voir photos ci-dessous). Ainsi, les ommatidies d’un même oeil composé recevant un même profil lumineux seront stimulées différemment. Ce mécanisme permet une vision polychromatique tout en n’ayant qu’un seul type d’opsine : en d’autres termes, des photorécepteurs identiques peuvent générer des sensibilités spectrales différentes.
Il n’y a pas d’images dans cette galerie.
Percevoir son environnement en couleur offre la possibilité d’interagir de manière plus complexe avec celui-ci : recherche de partenaire sexuel, phototaxie (déplacement en fonction de la lumière), oviposition (comportement de ponte), détection de prédateurs… Par exemple, des espèces d’hyménoptères, de lépidoptères ou de diptères ont pu être entrainées à associer des couleurs avec des comportements (vidéo ci-dessous). Aussi, il a été démontré chez des abeilles et des guêpes que les insectes possèdent également la capacité à discriminer une forme pas sa couleur.
De nombreuses espèces d’insectes privilégient certaines couleurs dans la recherche de nourriture, ce comportement pouvant être modifié par l’apprentissage et l’expérience. Par exemple, les insectes phytophages ne sont pas uniquement attirés par des sources lumineuses vertes, mais également par des sources jaunes. Les abeilles sont quant à elles attirées par des sources lumineuses de violettes à bleues, une réponse évolutive au fait que les fleurs reflétant de telles couleurs contiennent davantage de nectar.
IV. Discrimination du plan de vibration (sensibilité de polarisation)
Les ondes lumineuses vibrent dans un plan perpendiculaire à l’axe dans lequel elles se dirigent (illustration 5 ci-dessous). Ces plans de vibration peuvent être distribués sur les 360° entourant l’axe de direction. Lorsque toutes les ondes lumineuses vibrent dans un plan spécifique, la lumière est dite polarisée.
Illustration 5 : Plan de vibration et de polarisation de la lumière (Source : meteosat.pessac)
Illustration 6 : La lumière du soleil se polarise horizontalement dans un plan vertical (ici en gris) et verticalement sur un plan horizontale (ici en rose). Ainsi, en fonction de l’angle formé entre l’observateur et le soleil, un profil spécifique de lumière polarisée est créé. L’insecte utilise ce profil pour s’orienter, même lorsque le soleil n’est pas visible (Source : gatinel.com)
Par exemple, la lumière bleue provenant du ciel est polarisée : le degré de polarisation et le plan de polarisation maximale dépendent de la position du soleil, et ne sont donc pas les même dans les différentes parties du ciel (illustration 6 ci-contre). La détection de ces paramètres permet de déterminer la position du soleil, même lorsque celui-ci n’est pas visible. Certains insectes utilisent ces informations pour se diriger, notamment les insectes sociaux comme les abeilles (genre Apis) ou les fourmis du genre Cataglyphis qui recherchent de la nourriture puis reviennent à la colonie. Chez des insectes comme les Odonates (libellules) et les Diptères (mouches), la capacité à percevoir la lumière polarisée permet de maintenir une navigation stable et constante.
La perception du plan de polarisation est possible grâce à la présence de molécules pigmentaires photosensibles orientées dans un axe spécifique et situées dans les microvilli des rhabdomères. L’activation survient (absorption maximale) lorsque la lumière vibre dans le même plan que ces molécules (Illustration 7 ci-dessous).
Illustration 7 : Les molécules pigmentaires sont activées lorsque le plan de polarisation de la lumière leur est parallèle, pour s’orienter, l’insecte se place de sorte que la majorité des molécules pigmentaires soient parallèles au plan de polarisation de l’environnement (Source : gatinel.com) (Modifié par B. Gilles)
Dans les ommatidies, le rhabdomère est le plus souvent courbé (forme convexe de l’oeil composé), il ne peut donc y avoir de plan, d’absorption spécifique. La sensibilité de polarisation dépendra donc d’une orientation uniforme des molécules de pigments visuels dans un rhabdomère. Chez les fourmis et les abeilles, ainsi que chez d’autres insectes sensibles au plan de polarisation, des rhabdomères droits sont présents uniquement dans un petit groupe d’ommatidies situé le long du bord dorsal de chaque oeil. Ces ommatidies constituent à peine 6% du total des ommatidies chez la fourmis Cataglyphis et 2,5% chez les abeille du genre Apis. La stimulation des cellules photoréceptrices a lieu lorsqu’une majorité de molécules pigmentaires sont parallèles au plan de polarisation.
Le fonctionnement de ce système demeure encore lacunaire. L’hypothèse suggère qu’en balayant le ciel, l’insecte est capable de faire correspondre ces récepteurs de polarisations aux plans de polarisation du ciel. Lors de la réorientation, le motif du ciel correspondrait à l’entrée mémorisée. Une carte mentale pouvant réaliser ce genre de comparaison a été localisée dans le corps central du cerveau de criquets acridiens.
Source :
– The Insects : Structure & Function (5ème édition, 2013) – Edition : S.J. Simpson & A.E. Douglas – Cambridge University Press
– Encyclopedia of Insects : Vincent H. RESH & Ring T. CARDE, Academic Press-2009
– Tibbetts E. A. & Izzo A. (2010) : Social punishment of dishonest signalers caused by mismatch between signal and behavior – Current Biology, 20(18):1637-1640 (lien)
Recommandation d’ouvrages sur cette thématique
– Evolution of the Insects(David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
– The Insects: Structure and Function (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
– Encyclopedia of Insects (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
Avec la publication de son ouvrage, « Moi, parasite« , Pierre Kerner offre enfin une tribune à des organismes trop souvent méprisés au seul motif qu’ils vivent aux dépens d’autres espèces. Partez à la rencontre d’une multitude de parasites en tout genre, et notamment d’insectes. Ces histoires ne laisseront personne indifférent et raviront les curieux en quête de sensations fortes : entre les vers qui poussent des insectes à se noyer et s’extirpent par leur anus, les tiques qui rendent végétariens ou encore les bébés moules qui vampirisent les poissons, notre monde repousse toujours plus loin les limites de l’extraordinaire et du répugnant !
Interview de Pierre Kerner
Auteur de l’ouvrage : « Moi, parasite » (Mars 2018)
Maître de Conférence – Paris Diderot
Bonjour Pierre, peux-tu te présenter rapidement, pas ton toi parasite, mais ton toi humain ?
Pierre Kerner durant une conférence (Source : Lyon Science)
Mon moi humain est Enseignant Chercheur en Génétique Evolutive du Développement à l’Université de Paris Diderot où j’enseigne l’Evolution, la Zoologie, le Développement et bien entendu, un peu de Parasitologie. Mes recherches concernent l’évolution des animaux, et plus particulièrement celle des cellules souches que j’étudie chez un ver marin : Platynereis dumerilii. Depuis 2009, je suis blogueur sur Strange Stuff and Funky Things où je parle de sujet surprenants, essentiellement en Biologie. Je suis membre actif de la communauté de vulgarisateurs du Café des Sciences au sein de laquelle j’ai lancé des projets comme Strip Science (une plateforme tournée vers la rencontre entre illustration et science) et Vidéosciences (rebelote mais entre la vidéo et la science). Je sévis aussi régulièrement sur Podcast Science, un balado francophone scientifique amateur, et j’ai lancé ma chaine Youtube il y a maintenant 2 ans. Tel un parasite, je me suis insinué dans toutes les anfractuosités de la vulgarisation scientifique !
Tu viens de publier un ouvrage, « Moi, parasite« , où tu présentes la vie secrète de multiples parasites, comment l’idée d’aborder un tel sujet a-t-elle parasité ton esprit ?
Sur mon blog, j’ai abordé très rapidement le sujet du parasitisme (Catégorie Freaky Friday Parasite), tant le sujet est prolifique en exemples sordides, surprenants et fascinants. Je me suis essentiellement inspiré des billets de blogs de Carl Zimmer, Ed Yong et Christie Wilcox, mais aussi de mes propres découvertes lors de mes pérégrinations sur le web. Quand j’ai été titularisé en tant que Maître de Conférences, j’ai eu l’opportunité de reprendre des cours en Master sur la parasitologie : je me suis rué dessus et j’ai commencé à faire plein de recherches pour préparer un enseignement de qualité, tout en insérant les trouvailles que j’avais déjà bloguées. J’aime bien ces contributions à double sens entre mes enseignements et mes productions de vulgarisation. Que ce soit en cours ou en billets de blog, j’ai vite constaté que le sujet fascine et que le parasitisme permet d’aborder des aspects fondamentaux de la biologie tout en retournant plein d’idées reçues qu’on peut avoir sur la nature. Quand on m’a demandé quel livre je voudrais écrire, j’ai automatiquement pensé au sujet du parasitisme.
Tu abordes la biologie, l’écologie et l’évolution des relations hôtes-parasites sous un angle original, celui de la première personne, pourquoi se mettre à la place du parasite, un fantasme ?
Non, un stratagème d’écriture. Quand j’ai commencé le projet du livre sur le parasitisme, en discutant avec les deux illustrateurs Adrien Demilly et Alain Prunier et avec Laurent Brasier, le Responsable Editorial de la collection Science à Plumes de Belin, on a vite écarté la possibilité de faire un simple copier-coller de mes articles de blog. D’une part, j’avais envie d’aborder les aspects que je développais dans mes cours de Parasitologie et qui sont assez complexes, et d’autre part, je voulais que les histoires aient un liant entre elles. En brainstormant, on est arrivé avec l’idée d’incarner les parasites. J’ai rigolé en pensant qu’on pourrait titrer le livre « Moi, parasite » en référence au « Moi, Président » de François Hollande… Comme quoi, ça a du bon de rigoler. Au fur et mesure du processus d’écriture, je me suis rendu compte qu’incarner les parasites facilitait le côté emphatique, et, même si c’était une contrainte de taille, permettait aussi d’aborder des sujets sur le parasitisme depuis un angle neuf, même pour les parasitologues chevronnés ou les amateurs de ce genre d’histoires.
Le parasitisme est un sujet qui te passionne depuis toujours, ou l’as-tu découvert par la suite ? Qu’est-ce qui te fascine chez les parasites ?
J’ai vraiment nourri un intérêt pour le parasitisme avec mon blog. Au début, je cherchais à écrire des billets de blog sur des sujets surprenants, et le parasitisme revenant souvent parmi les exemples exploitables. Le fait est que les parasites exercent une fascination morbide pour nous : on est à la fois dégoûté mais aussi subjugué par leur mode de fonctionnement, leur cycle de vie complexe, leurs stratégies pour survivre, pour puiser des ressources. C’est comme un attrait pour les films d’horreur : on est des amateurs de sensations fortes et les parasites ne laissent pas de marbre. Par contre, ce qui est particulièrement agréable avec le parasitisme, c’est la propension que ce sujet a de pouvoir dissiper les idées reçues et repousser les limites du concevable. Les parasites sont d’excellents exemples de bricolages évolutifs incroyables.
Parmi les nombreux types et exemples de parasitismes que tu listes dans l’ouvrage, quel est celui dans lequel tu aimerais te réincarner dans ta prochaine vie ?
Aussi surprenant que cela puisse paraître, j’aime écrire sur les parasites, mais je ne suis pas particulièrement attiré par eux, ni par leur mode de vie. Comme tout un chacun, je frémis à l’idée de pouvoir être l’hôte de parasites. Une des raisons qui me pousse à écrire sur les parasites, c’est de pouvoir justement un peu exorciser mes peurs. Du coup, les parasites, je les respecte de plus en plus… Mais de là à vouloir en être un…
Le monde des parasites fait peu l’objet de communication auprès du grand public, il demeure mystérieux et inconnu, pourquoi ?
Suivez « Moi, parasite » sur les réseaux sociaux
Je ne sais pas si je suis foncièrement d’accord avec cette idée. C’est probablement un effet de bulle médiatique, mais au contraire, j’ai l’impression qu’on parle de plus en plus du parasitisme. Quand en 2009 j’apprenais l’existence de champignons parasites contrôlant le comportement de fourmis, en effet, très peu de personnes en parlaient. Mais aujourd’hui, c’est un exemple que beaucoup de personnes connaissent, notamment du fait de l’adaptation qui en a été faite dans le jeu vidéo The Last of Us, ou les nombreux billets de blogs et les vidéos Youtube qui évoquent ce phénomène. Un des défis du livre a été d’ailleurs de trouver des exemples originaux de parasitisme par rapport à ceux qu’on trouve assez régulièrement sur internet. Mais c’est peut être un effet de niche.
Pour toi, reste-il encore beaucoup de choses à découvrir sur les parasites en général et ceux parasitant les êtres humains ?
Pendant mes recherches lors de l’écriture de « Moi, parasite« , j’ai intégré des découvertes réalisées en 2017 et en 2018, quelques semaines avant la sortie du livre : le domaine de la parasitologie est vaste et des découvertes sont réalisées périodiquement. Partant du constat que la plus grande majorité des formes de vie pratiquent le parasitisme, il n’est pas étonnant qu’il y ait encore tellement de chose à découvrir sur eux ! Rien que dans le domaine de la taxonomie, lorsqu’on découvre une nouvelle espèce, il faut bien s’imaginer qu’on a en réalité souvent découvert un hôte de nombreuses espèces parasitaires en même temps, qu’il va falloir étudier et décrire.
Enfin, aurais-tu une anecdote de parasite des plus ragoûtantes à nous raconter ?
Ver Phasmarhabditis hermaphrodita parasitant une limace (Source : Wikipedia)
En faisant des recherches sur les parasites employés par des humains pour les assister dans leur lutte biologie contre des nuisibles, je suis tombé sur le mode de fonctionnement de nématodes de l’espèce, Phasmarhabditis hermaphrodita, qui sont capables de tuer leurs hôtes : des limaces (on parle alors d’espèces parasitoïdes). Mais, prouesse, ils sont en mesure de contrôler le comportement de leur hôte et de pousser les limaces parasitées à s’enterrer avant de mourrir pour éviter que les larves de nématodes se retrouvent à l’air libre… Pratique quand on est un cultivateur et qu’on ne souhaite pas voir son potager couvert de cadavres de limaces !
Où se procurer l’ouvrage :
– « Moi, parasite » – Pierre Kerner – Editions Belin – Collection Sciences à plumes (192 pages – 14 mars 2018)
Dans la même rubrique, vous pouvez découvrir les interviews de :
David GIRON (entomologiste-chercheur CNRS – IRBI-Université de Tours)
Dans le monde des fourmis à la diversité déjà surprenante : morphologie, comportement, adaptation et écologie (lire ces articles : Cataglyphis ; Atta ; Eciton ; Cephalotes ; Force), s’ajoute la description d’une nouvelle espèce à la capacité des plus singulières, celle de pouvoir se faire littéralement exploser comme moyen de défense ! Décrite en ce début d’année par deux entomologistes autrichiens, Alice Laciny et Herbertt Zettel, l’espèce a été logiquement nommée : Colobopsis explodens.
Classification
Cette nouvelle espèce rencontrée dans les forêts tropicales de Bornéo, de Thaïlande et de Malaisie appartient à un groupe d’espèces monophylétiques, celui de Colobopsis cylindrica (COCY) ou de C. saundersi (Emery, 1889). Ce groupe est également appelé vernaculairement « fourmis explosives » en raison de la capacité des ouvrières à se sacrifier durant des combats territoriaux en faisant volontairement exploser par rupture des téguments de leur ganter (autothysis) et en relâchant des éléments irritants et gluants par leurs glandes mandibulaires hypertrophiées pour tuer ou pour repousser l’ennemi.
Le groupe COCY fait l’objet depuis une dizaine d’années d’études écologiques, morphologiques et chimiques. En 2014, une équipe pluridisciplinaire a débuté un programme de recherche pour explorer l’évolution de l’autothysis. Pour cela, les scientifiques ont étudié 15 espèces au Kuala Belalong Fields Studies Center (KBFSC) dans le Brunnei. C’est dans ce contexte que l’analyse moléculaire et taxonomique d’une espèce arborant des glandes sécrétrices mandibulaires de couleur jaune brillant – découverte près du centre KBFSC – a révélé qu’il s’agissait d’un morphotype (traits morphologiques spécifiques) d’une espèce non décrite : Colobopsis explodens.
Les chercheurs se sont intéressés à cette fourmi un peu par hazard : elle possédait un comportement défensif plus prononcé et était la plus abondante en périphérie du centre.
Description morphologiques
Morphotype d’une ouvrière minor de C. explodens : a) vue de face – b) vue latérale – c) vue dorsaleMorphotype d’une ouvrière major de C. explodens : a) vue de face – b) vue frontale – c) latérale – d) vue dorsaleMorphotype d’une reine de C. explodens : a) vue de face – b) vue frontale – c) vue latérale – d) vue dorsale – e) aile antérieureMorphotype d’un mâle de C. explodens : a) vue de face – b) ailes antérieure et postérieure – c) vue latérale – d) vue dorsale
Biologie & écologie
Les colonies de C. explodens ont été découvertes dans le Ulu Temburong National Park (carte ci-contre). Ces fourmis sont polygones (plusieurs reines au sein d’une colonie) et polydômes (réparties entre plusieurs nids). Les nids de C. explodens, pouvant abriter plusieurs milliers d’individus, se rencontrent le plus souvent dans des arbres de la famille des Dipterocarpaceae (regroupant 70 espèces en Asie).
Le réseau de nids étudiés s’étendait à la fois dans la canopée, entre 25 et 60m de hauteur, et sur le sol, entre plusieurs espèces d’arbres sur une superficie estimée à près de 2 500 m2. Pour l’expérimentation, 4 d’entre eux ont été prélevés puis installés dans des nids artificiels durant plusieurs semaines. Il a pu être mis en évidence une activité diurne de recherche de nourriture (foraging), principalement entre 9h et 16h lorsque la température est la plus élevée (24-28,6°C).
Une colonie de C. explodens se compose de plusieurs castes d’ouvrières, où les formes minor (les plus petites fourmis) sont les plus abondantes et les formes major, les moins nombreuses, ne sortent quasi-jamais du nid. C’est la caste des minor qui se sacrifie en libérant un liquide gluant de couleur jaune d’odeur épicée.
L’alimentation de ces fourmis demeure en partie inconnue. Il a été observé, in natura, des minor passant de longs moments à patrouiller et débarrasser la surface des feuilles de nombreuses débris et autres petits arthropodes, et à « brouter » sur les épiphytes (mousses, lichens, algues, filaments fongiques) avec leurs mandibules, parfois pendant près d’une heure. Ces comportements sont peut-être liés à la nutrition des fourmis. In vitro, les fourmis ont accepté de consommer de petits insectes morts et des fruits que leur proposaient les scientifiques.
L’analyse génétique du contenu des glandes sécrétrices mandibulaires de minor Colobopsis explodens a révélé la présence de bactéries du genre Blochmania sp. (Gammaproteobactéries), des symbiotes trouvés chez les fourmis Camponotus sp. (dites charpentières).
Les scientifiques indiquent que la description du mécanisme de l’autothysis, la composition des glandes sécrétrices mandibulaires, l’association micro-organismes/COCY et la nutrition de C. explodens feront l’objet de prochaines publications…
Affaire à suivre !!
Source :
Laciny A. ; Zettel H. ; et al. (2018) : Colobopsis explodens, model species for studies on « exploding ants », with biological notes and first illustrations of males of the Colobopsis cylindrica group. Zookeys 751:1 1-40 (lien)
Passionné depuis l’enfance par les « choses de la nature », j’opte pour un bac scientifique appelé à l’époque « Moderne prime ». Ce premier diplôme en poche, je pars à la recherche d’une formation où l’eau aurait un rôle principal, avec en tête l’idée de faire de la pisciculture à caractère plus scientifique qu’un simple élevage intensif de poissons. Mes regards se dirigent vers une formation « Eaux-et-Forêts », quand je découvre par hasard que l’ENSAT (Ecole Nationale d’Agronomie de Toulouse) propose une formation spécialisée en pisciculture et biologie des eaux…
Parti pour la « Ville Rose » pour une année de préparation, j’intègre cette école mais me rends vite compte que cette spécialité annoncée y est très mal enseignée, voire sans intérêt. J’y suis… j’y reste, mais un peu désabusé et, ayant la possibilité de suivre en même temps un enseignement en faculté, je vise en parallèle l’obtention d’une licence en Biologie à la Faculté des Sciences de Toulouse.
Beaucoup de travail à la clef, mais aussi le bénéfice de me frotter à deux enseignement différents, l’un très pratique en Ecole d’ingénieur et l’autre très théorique et cumulatif en Université. Ces deux manières d’acquérir des connaissances me seront longtemps bénéfiques par la suite.
Claude Dejoux étudiant les insectes aquatiques du lac Tchad (Source : C. Dejoux)
Obtenant une licence de Biologie et un diplôme d’Agronomie, j’ai mon passeport pour une carrière de recherche en écologie, avec comme outils de départ mathématiques, physique, chimie et biologie générale, dont un certificat de licence spécialisée en entomologie ! Apprenant par hasard que l’Office de la Recherche Scientifique et Technique d’Outre-mer (ORSTOM) (devenu l’IRD) recrute des hydrobiologistes, je postule par une simple lettre et suis immédiatement accepté comme élève chercheur, une douce époque malheureusement révolue !
Après un an et demi de complément de formation dans différents laboratoires (Paris, Banyuls en Océanographie, Biarritz et le CNRS de Gif-sur-Yvette en Hydrobiologie, Institut Max Plank de Plön en Allemagne pour une spécialisation sur l’étude des Diptères Chironomidae et à Hambourg sur les problèmes de pollution des eaux), je fais trois mois de « classes » en France, avant de partir terminer mon service militaire, en 1964, comme VSN (Volontaires du Service National) au Tchad, où je resterai travailler une dizaine d’années dans le cadre de l’ORSTOM.
Ayant au départ choisi la composante insectes aquatiques au moment de la constitution par l’ORSTOM de son équipe d’hydrobiologistes au Tchad, je travaille tout d’abord sur le Lac Tchad, puis sur d’autres milieux aquatiques du pays. Taxonomie et écologie des Chironomidae occupent la majorité de mon temps dans les premières années, vu leur importance dans la chaine alimentaire des poissons et des oiseaux de cet écosystème, travaux qui déboucheront sur une thèse d’Etat soutenue à l’Université d’Orsay.
Larves de simulies (Simulium damnosum) sur un support végétal, en eau courante (Source : C. Dejoux)
Le projet de mise en place par l’OMS (Organisation Mondiale de la Santé) d’un long et vaste programme de lutte contre l’onchocercose, filariose transmise par une simulie (petit Diptère dont les larves vivent dans les eaux courantes des rivières et fleuves d’Afrique de l’Ouest) est pour moi une opportunité d’appliquer mes connaissances sur les insectes aquatiques. En effet, le programme vise à une éradication de ce Diptère dans les principaux cours d’eau par application hebdomadaire d’insecticides sur les gîtes larvaires, durant une vingtaine d’années, tout en préservant les autres espèces aquatiques non cibles.
Quittant le Tchad pour la Côte-d’Ivoire, base opérationnelle plus centrale pour ce programme, je mets au point plusieurs systèmes de tests de toxicité des insecticides « candidats », tant en laboratoire qu’in situ dans les cours d’eau. Ces méthodes de recherche d’impact seront utilisées et affinées durant les 7 années que durera ma collaboration à ce programme qui s’avérera finalement un succès. Sensibiliser depuis plusieurs années sur les problèmes d’atteinte des milieux aquatiques par les pesticides et toutes autres causes de pollution, je suis mandaté en 1984 par le programme international « GEMS Water » (Programme de Surveillance Mondiale de la Qualité de l’Eau, PNUE-MOS-UNESCO-OMM) afin de rechercher dans toute l’Afrique de l’Ouest des laboratoires de référence pouvant réaliser des prélèvements réguliers et des analyses alimentant la base de données de cette surveillance mondiale. Cela m’amènera à la rédaction d’un travail de synthèse relatif à la pollution des eaux continentales africaines publié en 1988 (Editions ORSTOM).
Ayant formé suffisamment de techniciens locaux pour continuer un travail devenu routine, j’accepte la proposition de l’ORSTOM pour prendre en charge la représentation de cet institut à La Paz, en Bolivie. La tâche administrative est lourde mais je trouve l’occasion d’étudier l’entomofaune de la partie bolivienne du Lac Titicaca et, avec un collègue algologue, de réaliser une synthèse des connaissances limnologiques de ce magnifique milieu d’altitude. Cette synthèse sera publiée en 1992 (Editions Kluwer) dans la série des monographies biologiques des grands lacs du monde.
Pollution de l’eau à Chalco, quartier périphérique de la ville de Mexico (Source : C. Dejoux)
Après 4 années (1985-1989) passées à travailler entre 3 500 et 5 000 mètres d’altitude, je redescends un peu en acceptant le poste de Représentant de l’IRD à Mexico en 1992, où, à une altitude idéale de 2 400m, il me faut quand même partager l’oxygène avec quelques 20 millions de concitoyens ! J’y finirai une carrière devenue administrative, mais faite de coordination de programmes multidisciplinaires qui me donneront une vision écologique (au sens étymologique du terme…) des problèmes liés à la préservation de différents types d’environnement de notre planète. Cette vision d’interdépendance des facteurs écologiques va, une fois à la retraite en 1996, me permettre des interventions variées sur des problèmes environnementaux dans plusieurs pays du monde par le biais de l’ECTI, une association de séniors bénévoles.
Importance des insectes dans les milieux aquatiques
Alors que pratiquement absent des océans (une seule famille d’Hémiptères semi-aquatiques s’y rencontre : Halobatidae), les insectes peuplent tous les milieux d’eau douce avec dans certains une très grande diversité.
Dans la majorité des cas, ils y passent leur phase larvaire et nymphale (Odonates, Ephémères, Diptères, etc.), la phase adulte étant aérienne, permettant dissémination et reproduction. Véritable microcosme (bien que certaines espèces atteignent plusieurs centimètres), ils ont un rôle important dans les équilibres des biotopes aquatiques en participant à la dégradation de la matière organique comme décomposeurs primaires (par fragmentation, ingestion et défécation), avant l’action des décomposeurs secondaires, vers et bactéries par exemple.
Baterie de gouttières en PVC permettant de tester simultanément plusieurs insecticides (Source : C. Dejoux)
Leur rôle est tout aussi important dans la chaine alimentaire, servant de nourriture à de nombreux poissons, batraciens et oiseaux après avoir eux même consommés des micro algues ou des macrophytes. Le bol alimentaire de certains poissons africains comme les Mormyridae peut être exclusivement composé de larves de Diptères Chironomidae (« vers de vase rouge » des pêcheurs), comme celui de quelques Anatidae (canards) se nourrissant sur les vasières de faible profondeur. Ne parlons pas de toutes les espèces opportunistes : poissons, batraciens, oiseaux qui se gavent par exemple lors d’émergences en masse d’Ephémères.
Pénétrer l’environnement des insectes aquatiques, c’est aussi s’étonner des adaptations qu’ils ont su développer pour coloniser toutes sortes de micro-milieux et, pour chaque espèce, rechercher son preferendum environnemental. Comprendre pourquoi une espèce est ou n’est pas présente dans un milieu est la clef des études d’écologie dynamique et aide mieux à analyser les impacts des aggressions des hydrosystèmes.
Les insectes, bio-indicateurs de pollution
Si l’on prend le terme pollution dans son sens le plus large, i.e. une agression du milieu aquatique avec impact négatif sur son intégrité, les insectes constituent un très bon outil d’analyse. Ils ont chacun, comme chaque être vivant, leurs faiblesses, tant physiques que physiologiques et, selon la nature d’une pollution et la connaissance de l’écologie que l’on peut appeler « intime » des espèces présentes, il est possible de déterminer celles qui seront le plus affectées.
Tests de toxicité entre deux insecticides in situ (Source : C. Dejoux)
De nombreux indices ont été proposés par différents auteurs afin de quantifier un niveau de pollution ou de salubrité d’un biotope aquatique : beaucoup utilisent les insectes dans leur calcul. Pour l’études des cours d’eau par exemple, l’indice le plus fréquemment utilisé en France est l‘IBGN (Indice Biologique Global Normalisé) qui, en fonction des densités relatives de certaines familles d’insectes (Trichoptères, Ephéméroptères, etc. ), aboutit à une note finale qui se veut caractériser le niveau d’agression du milieu. Ces méthodes sont rapides mais traduisent mal l’action des polluants et ne peuvent prendre en compte certaines perturbations naturelles survenues quelque temps auparavant et dont l’effet peut affecter à moyen terme l’hydro-système. Une bonne connaissance générale et sur une longue durée de la dynamique des milieux étudiés permet de pondérer le résultat des analyses ponctuelles.
C’est certainement en combinant ces deux types de connaissances que nous sommes parvenus, dans les rivières africaines traitées aux insecticides anti-simulidiens, à évaluer l’impact à long terme sur les entomocénoses (communautés d’êtres vivants interdépendants, ici des insectes, occupant un même biotope). Cet impact s’est avéré plutôt faible avec le Téméphos (insecticide organe-phosphoré) longtemps utilisé, plus important avec le Chlorphoxim (organe-chloré) introduit par la suite pour éviter de développer une résistance croisée chez les insectes cibles et pratiquement nul avec l’utilisation de Bacillus thuringiensis (BTI, souche H14-De Barjac). Parmi les familles les plus sensibles aux deux premiers composés, il faut citer les Ephémères de la famille des Baetidae ainsi que les Plécoptères Perlidae.
Pollution des milieux aquatiques dans différents pays et pollution environnementale générale
Les données initiales (temps, température, etc. à une certaine date) manquent encore souvent pour statuer avec précision sur l’évolution de la pollution des écosystèmes aquatiques mondiaux (même si c’était le but du programme GEMS Water).
Ma propre expérience me fait dire que la qualité des eaux continentales est globalement meilleure dans de nombreux pays, et de plus en plus préoccupante dans d’autres ! Elle est meilleure quand des législations adaptées régissent les rejets polluants dans les milieux aquatiques, voire même la plus simple utilisation de l’eau douce naturelle. Elle est pire quand rien ne protège ces milieux ou quand la législation n’est pas appliquée.
Récolte de la faune benthique du lac Titicaca à l’aide d’une benne d’Ekman (Source : C. Dejoux)
Dans de très nombreux pays, l’homme considère les cours d’eau comme de simples égouts, capables à la fois de tout digérer sans conséquences et surtout de transporter les déchets… ailleurs ! L’augmentation permanent de la population mondiale, liée à l’avidité de profits sont deux facteurs extrêmement préoccupants. Cours d’eau canalisés, urbanisme galopant, déforestation, usage démesuré de pesticides, agriculture et industries avides d’eau sont autant de facteurs qui affectent les milieux aquatiques, même si la présence de stations d’épuration (quand elles existent) soit un facteur positif et donnent en plus bonne conscience.
Sans plagier Arthus Bertrand ou Nicolas Hulot, j’ai eu par exemple l’occasion de voir en Chine les cours d’eau pollués par les ruissellements de pesticides d’une agriculture intensive, les résidus miniers divers ou les accidents « écologiques graves » qui détruisent toute vie aquatique durant de longs mois, des fleuves transportent des millions de mètres cubes de sédiments, suite à l’érosion des bassins versants due à l’appauvrissement du couvert végétal. Au Maroc, l’eau de boisson de Rabat est difficile à traiter en raison de l’eutrophisation des lacs d’approvisionnement, phénomène lié au lessivage des terres recevant trop de fertilisants. En Bolivie, une usine de textiles déversant directement ses « effluents colorés » dans le Rio la Paz… devenu azoïque sur des kilomètres ou bien les effluents d’une mine de fer directement déversés dans un lac dont le pH était descendu à 1,5. Au Mexique, des rejets des eaux usées de la ville de Mexico provoquant dans les cours d’eau les recevant des montagnes de mousse issues de l’utilisation abusive de détergents domestiques…
Tout ceci ne concerne que l’eau, mais il est possible de faire des constats semblables concernant la terre ou l’air et il n’y a qu’à se pencher sur les taux de pollution dans les grandes villes ou de constater la dégradation des sols naturels en raison d’une agriculture intensive non raisonnée, la perte d’une couverture végétales stables entraînant érosion, ruissellement et pertes de matière organique…
Je pense que la prise de conscience existe, mais reste insuffisante. Un effort d’information plus important s’impose auprès des jeunes et l’écologie, au sens noble du terme, devrait avoir une place de choix dans les programmes scolaires.
Evolution des atteintes à l’environnement dans le monde
Bien qu’il soit nécessaire de distinguer les situations en fonction du degré de développement des pays concernés, je pense qu’un peu partout il est possible de distinguer deux phénomènes dont les résultantes s’opposent. D’une part l’augmentation presque exponentielle de la population dans le monde crée de plus en plus de besoins pour nourrir et assurer la vie au sens large de toutes ces personnes. Cela implique une exploitation de plus en plus grande et parfois (souvent ?) anarchique de toutes les resources naturelles, avec les conséquences que l’on sait sur l’environnement : déforestation, épuisement des énergies fossiles, agriculture intensive épuisant les sols (ou extensive réduisant la biodiversité…), destruction des habitats naturels pour le simple profit, la liste des impacts de cette humanité grandissante est longue et nous ne listons là que quelques macro phénomènes .
En opposition, de nombreux scientifiques ou simples citoyens ont pris conscience de cette situation et s’efforcent de la freiner ou de trouver des solutions de remplacement. Les connaissance en écologie ont énormément progressé et il n’est plus nécessaire de voir des poissons le ventre en l’air dans un cours d’eau pour se rendre compte si le milieu est pollué. De même, on réalise maintenant que l’utilisation trop intense de l’irrigation épuise les nappes plus vite qu’elles se rechargent et surtout diminue par lessivage la fertilité naturelle des sols.
Un peu partout des mouvements se créent pour le respect de la nature, la préservation d’une biodiversité dont le degré, élevé ou faible, reflète en partie la bonne ou la mauvaise santé de notre planète. L’agro-écologie prouve que l’on peut nourrir beaucoup de gens sans pour cela cultiver des salades sans sol dans des hangars gigantesques ! Le recyclage, au sens large du terme, devient lentement une philosophie économique aux effets bénéfiques (voir l’article sur l’entomoculture).
Région par région, la situation rencontrée à l’heure actuelle sera une résultante de ces deux grandes tendances, mais je pense qu’il faudra encore du temps pour que nous prenions tous conscience de la nécessité de respecter le plus possible cet énorme potentiel qui nous entoure : l’eau, l’air et la terre !
Trop souvent encore, le verbe avoir est préféré au verbe être et le mot « profit » gouverne nos comportements. J’ai par exemple réalisé deux missions en Mongolie intérieure chinoise, au même endroit, à 7 années d’intervalle. La première avait pour but la préservation et la restauration des zones humides au nord du massif tibétain. La seconde, d’écologie plus générale, était liée à la réussite du programme national de reforestation du nord de la Chine pour ralentir la désertification.
Mon rapport de première mission préconisait des mesures de conservation des zones humides existantes et surtout sonnait le signal d’alarme devant une utilisation anarchique des ressources en eau locales. Sept années plus tard, la ville qui m’accueillait était passée d’une population de 300 000 habitants à plus d’un million. Des buildings gigantesques poussaient de tout coté, de beaux parcs abondamment arrosés ornaient les quartiers résidentiels et, cerise le gâteau, un grand lac artificiel « récréatif » avait été créé, alimenté par des pompes prenant l’eau dans le fleuve proche qui n’était plus qu’un « squelette » de cours d’eau. A coté de cela, toute la vallée fluviale en aval de la ville, où existait 7 ans plus tôt une agriculture florissante, était devenue quasiment désertique et pour nourrir des milliers d’habitants supplémentaires, toutes les denrées vitales étaient importées du sud de la Chine. Ne parlons pas en plus des milliers d’arbres morts au niveau de l’implantation de la fameuse « ceinture verte », par défaut d’apports minimums en eau !
Je pourrais ainsi multiplier ces exemples d’abrasion environnementale comme l’assèchement presque total d’une partie du cours de la Volta pour arroser le périmètre sucrier de Banfora au Burkina Faso, ou les cultures du coton en zone sahélienne qui ont dévasté la brousse au détriment des cultures vivrières pour finalement aboutir à une production pratiquement sans valeur sur le marché mondial. En Bolivie, le lac Titicaca qui était plein de vie dans sa région sud est dans les années 1980 est devenu dans cette région une soupe d’algues sans poissons par hyper eutrophisation, en raison de l’accumulation des nutriments provenant du lessivage des terres agricoles de bordure.
Bien entendu il ne faut pas voir que ce mauvais coté de notre agression des milieux naturels car de belles actions existent par ailleurs, mais les signaler aide à une prise de de conscience de nos responsabilités vis à vis de notre milieu de vie.
Une anecdote particulière…
Hélicoptère de prospection hydrobiologique du programme OMS de lutte contre l’onchocercose (Source : C. Dejoux)
Ayant rédigé un rapport défavorable à l’utilisation d’un insecticide proposé par les traitements de longue durée par la firme Bayer, ce travail a été contesté et pour prouver l’innocuité du produit un « expert » fut envoyé en Côte d’ivoire pour vérification. Avec lui nous avons refait des tests in situ avec des traitements grandeur nature, avec nos appareils mis au point pour ce faire et en comparant sur deux biefs différents les impacts de l’insecticide habituellement utilisé avec celui du nouveau proposé. Le résultat ayant été sensiblement le même : défavorable au nouveau produit, l’argumentaire de l’expert a été que nous aurions du faire le test en même temps et dans une zone de rapides, avec sur une moitié du cours d’eau un traitement avec un produit et sur la moitié symétrique avec le produit Bayer ! Quand on connait les turbulences liées aux zones de rapides, on imagine mal couper le bief dans le sens de la longueur pour que chaque produit reste de son coté ! Nous n’avons d’ailleurs jamais eu de nouvelles de l’expert et de son éventuel rapport.
Album photos
Dans la même rubrique, vous pouvez découvrir les interviews de :
David GIRON (entomologiste-chercheur CNRS – IRBI-Université de Tours)
La spécialisation se définit comme l’adaptation physique, biologique et/ou comportementale à un groupe restreint de partenaires. La bryone dioïque (Bryona dioica), une Cucurbitacée sauvage, fleurit au printemps dans les haies et les friches. Ses fleurs blanc verdâtre attirent divers butineurs. Parmi eux, une petite abeille noirâtre marquée de rouge, l’andrène de la bryone (Andrena florea), visite uniquement ses fleurs, malgré la masse d’autres plantes en fleurs en cette saison. De même en région méditerranéenne, le figuier (Ficus carica) n’est pollinisé que par une petite guêpe végétarienne (Blastophaga psenes) qui vient pondre dans ses inflorescences quasi fermées.
Sans le figuier, la guêpe ne peut survivre. Mais sans la guêpe, le figuier ne peut pas se reproduire correctement. Ces deux espèces ont tissées entre elles des relations obligatoires. Par contre si l’abeille de la bryone dépend totalement de cette plante pour sa survie, avec laquelle elle a donc développé une relation obligatoire, l’inverse n’est pas vrai. La bryone visitée et pollinisée par d’autres insectes n’entretient qu’une relation facultative avec son andrène.
Le généralisme, à l’inverse, se définit comme la faculté, ou la possibilité, de nouer des relations facultatives avec un grand nombre de partenaires. Parmi les insectes pollinisateurs, l’abeille mellifère (Apis mellifera) est un excellent exemple de butineur généraliste. Elle peut butiner des centaines d’espèces différentes de fleurs. Plantes autochtones ou plantes introduites, du moment que la fleur produit du nectar ou du pollen qui lui sont accessibles, elle peut venir les visiter. Ce généralisme a des limites, à cause notamment de la longueur modeste de sa langue, cinq à six millimètres. Les fleurs étroites où le nectar se trouve en profondeur lui sont inaccessibles.
Chez les plantes, le généralisme se traduit par la faculté de permettre à de très nombreux animaux d’accéder à leur nectar ou à leur pollen, comme chez les Ombellifères. Sur leurs ombelles groupant de nombreuses petites fleurs peu profondes se rencontrent la plupart des insectes floricoles, des coléoptères aux abeilles en passant par les mouches, les guêpes et les papillons. Mais ce buffet à volonté, en exacerbant la concurrence, fait fuir les espèces les mieux outillées qui préfèrent d’autres sources plus rentables. Les bourdons ou les papillons de jour, équipés d’une longue langue ou d’une trompe déroulante ne se voient quasiment jamais sur des Ombellifères.
L’auteur : Vincent Albouy
Vincent Albouy (Source : Christian Rome)
Entomologiste de formation, Vincent Albouy a présidé l’Office Pour les Insectes et leur Environnement (OPIE). Il s’intéresse depuis trente ans aux insectes communs et à leur protection, à la biodiversité ordinaire et au jardinage naturel qu’il pratique dans son jardin de Saintonge.
Auteur de nombreux ouvrages grand public sur la nature en général et les insectes en particulier, il vient de publier « Pollinisation : Le génie de la nature » aux éditions Quae.
Retrouvez les ouvrages de l’auteur en bas de page.
Le trouble des frontières
Cette distinction entre spécialistes et généralistes est une classification formelle qui n’a de réalité que dans l’esprit humain. Elle n’a aucune traduction concrète dans la nature où tous les cas s’observent en une lente gradation depuis les généralistes les plus larges jusqu’aux hyperspécialistes liés à un seul partenaire. D’ailleurs le généraliste parfait, capable d’attirer tous les animaux floricoles ou de butiner toutes les fleurs n’existe pas. Certains bourdons sont encore plus généralistes que l’abeille mellifère, mais uniquement actifs de jour ils ne peuvent visiter les fleurs qui ne s’ouvrent que la nuit. Et ce généralisme reste modeste. Le bourdon terrestre (Bombus terrestris), l’un des plus généralistes, a été noté comme butinant sur 309 espèces différentes de plantes dans nos régions, performance à mettre en relation avec les 4 500 espèces qui s’y rencontrent.
D’autre part, toutes les facettes de la biologie d’une espèce doivent être prises en compte. Les butineurs les plus étroitement spécialisés dans un aspect de leur comportement peuvent se révéler généralistes dans un autre. La mélitte de la lysimaque (Macropis europaea), une abeille solitaire, récolte exclusivement le pollen nécessaire à ses larves sur les fleurs de la lysimaque commune (Lysimachia vulgaris). C’est donc une hyperspécialiste pour le pollen car dépendante d’une seule espèce de plante. Mais la lysimaque ne produit pas de nectar. Alors la mélitte butine d’autres fleurs pour cette ressource, appartenant à des familles aussi diverses que celles de la marguerite, de la bourrache, de l’églantier. C’est donc une généraliste pour le nectar.
Abeille Macropis europaea butinant sur une fleur de Lysimachia vulgaris (Source : V. Albouy)
La situation se complique en observant finement les comportements de butinage au sein d’une même espèce. Deux individus peuvent avoir des choix de butinage très différents. Les études ont surtout concerné l’abeille mellifère, l’une des espèces les plus généralistes comme nous l’avons vu. La colonie exploite au mieux les ressources disponibles comme les analyses du pollen rentré à la ruche le prouvent : arbres, arbustes, plantes grimpantes, plantes herbacées, plantes sauvages ou cultivées, tout l’environnement dans un rayon de plusieurs kilomètres autour de la ruche est exploré et exploité.
Mais l’analyse du comportement des individus montre tout autre chose. La carrière des butineuses est courte au printemps et en été, au moment des grandes miellées, quand les floraisons et les ressources abondent. Deux semaines au plus. Et pendant ces deux semaines, la grande majorité des butineuses va se contenter d’exploiter quelques espèces seulement, voire une seule. Cette constance florale s’explique par une rationalisation du travail.
Butiner une fleur correctement et rapidement demande un peu d’apprentissage, surtout pour une espèce généraliste qui n’est pas préadaptée à la fleur qu’elle visite comme peut l’être une espèce spécialisée. Passer sans cesse d’un type de fleurs à l’autre fait perdre du temps et de l’énergie à la butineuse, donc des ressources à la colonie. D’où l’intérêt de ne se concentrer que sur une seule fleur, tant qu’elle est disponible et fournit des ressources alimentaires, pour gagner du temps et rentabiliser l’investissement dans l’apprentissage initial.
Abeille mellifère (Apis mellifera) butinant un tournesol (Source : V. Albouy)
Il en résulte que si l’abeille mellifère est une généraliste au niveau de l’espèce, elle peut se comporter comme une spécialiste au niveau individuel. Combien de butineuses ne connaissent que les fleurs de colza ou que celles de tournesol si elles appartiennent aux générations d’avril ou de juillet ? Seules environ 5% des butineuses, les éclaireuses qui repèrent les nouvelles ressources florales intéressantes et en informent leurs consœurs par une danse codée, se comportent vraiment en généralistes.
En résumé, un pollinisateur peut se comporter en généraliste ou en spécialiste selon le moment, le milieu, l’offre de fleurs à butiner, la présence ou non d’autres butineurs entrant en concurrence pour certaines ressources, etc.
Objectif pollinisation
Le concept de fleurs généralistes semble plus solide, du moins si l’on juge ce généralisme à l’aune du nombre de pollinisateurs animaux attirés. Une touffe de lierre en fleurs bourdonne d’une infinité d’insectes à l’automne, mouches, papillons, guêpes, abeilles. Son généralisme semble incontestable. Et pourtant c’est loin d’être évident si l’on raisonne en terme de pollinisation efficace.
Le seul but de la fleur, c’est d’être pollinisée, et correctement pollinisée. Donc du point de vue de la réussite de la reproduction d’une plante, il ne faut pas juger du nombre et de la variété de l’ensemble des visiteurs, mais seulement de ceux qui assurent une pollinisation efficace. Ces pollinisateurs efficaces peuvent se révéler au final peu nombreux.
Collette du lierre (Colletes hederae) sur des fleurs de lierre (Hedera helix) (Source : V. Albouy)
Le lierre (Hedera helix) dans nos régions est surtout pollinisé par l’abeille mellifère, butineur généraliste, et par la collete du lierre (Colletes hederae), une abeille solitaire très spécialisée puisque les femelles ne recueillent la nourriture des larves que sur le lierre. Dans le nord de l’Angleterre, où la Colletes hederae ne se trouve pas, les guêpes sociales prennent le relais et deviennent des pollinisatrices efficaces. Donc en jugeant le lierre à l’aune de sa pollinisation effective, il s’avère une espèce spécialisée sur les hyménoptères, et en particuliers sur les abeilles.
Beaucoup d’Ombellifères comme la carotte (Daucus carotta) attirent autant d’insectes que le lierre, et peuvent être pollinisées par la plupart d’entre eux : ce sont de vraies fleurs généralistes. D’où les concepts de « généralisme apparent « (le lierre) et de « généralisme fondamental » (les Ombellifères) introduits par les scientifiques. Mais une analyse fine des visiteurs de la carotte montre que ses pollinisateurs appartiennent à une catégorie bien précise : des insectes de petite taille à pièces buccales non spécialisées ou à langue courte. Certains la considèrent comme spécialisée sur cette catégorie de floricoles : coléoptères, mouches et guêpes principalement.
Syndrome de Pollinisation
L’existence de « syndromes de pollinisation » chez les fleurs est une idée largement admise par les botanistes. Ils entendent par cette expression l’ensemble des caractères d’une fleur adaptés au pollinisateur efficace de la plante, qui peut être le vent, l’eau ou des animaux comme les insectes, les oiseaux, les chauves-souris.
Ces caractère ou traits floraux, concernent aussi bien la taille, la forme, le type de symétrie de la fleur que sa couleur ou son odeur. Lorsque le pollinisateur est un animal, il présente en miroir des adaptations dans sa morphologie, sa physiologie, son comportement qui trahissent une co-évolution entre la plante et son pollinisateur, une relation de plus en plus étroite au fil du temps et de l’évolution sous la pression de la sélection naturelle. Traduite dans le langage courant, cette idée sous-entend qu’il existe des fleurs à abeilles, à papillons ou à oiseaux.
Exemple de co-évolution spécifique entre deux espèces : Colletes cunicularius et Orchidée Ophris occidentalis (La fleur mime la femelle de l’abeille par sa forme et par l’odeur pour attirer le mâle qui, en essayant de s’accoupler avec celle-ci, va féconder la fleur)
Les insectes ne voient pas le rouge, qui leur apparaît noir, alors que les oiseaux ont la même vision que nous. Le rouge est donc considéré comme caractéristique du syndrome de l’ornithophilie. En d’autres termes, une fleur rouge a de grandes chances d’être une fleur butinée par les oiseaux. Les mimules, les « fleurs singes » des anglophones, qui se trouvent surtout en Amérique du nord et en Australie ont donné une preuve expérimentale de cette affirmation.
Les espèces à fleurs rouges sont bien visitées et pollinisées par les oiseaux, et celles à fleurs roses le sont par les insectes. Pour confirmer cette adaptation de la couleur de la fleur à son pollinisateur, des généticiens ont manipulé la mimule de Lewis (Mimulus lewisii) rose pour la faire devenir rouge, et la mimule cardinale (Mimulus cardinalis) rouge pour la faire devenir rose. Cette simple modification de la couleur a entraîné des visites beaucoup plus importantes d’oiseaux sur la mimule à insectes devenue rouge et d’insectes sur la mimule à oiseaux devenue rose.
Certains biologistes mettent en doute la réalité des syndromes de pollinisation, en faisant remarquer qu’il est impossible de connaître les pollinisateurs efficaces d’une plante au simple examen de sa fleur. Trop d’exceptions à la règle des syndromes se présentent.
L’intérêt du compromis
En parallèle aux syndromes de pollinisation, la spécialisation des pollinisateurs doit aussi se juger sur des critères très larges, par exemple la possession d’une langue lécheuse longue (bourdons et certaines abeilles solitaires) ou d’une trompe aspirante (les papillons et certains bombyles). Les possesseurs d’un même type d’outillage constituent des « groupes fonctionnels ». Plusieurs espèces de butineurs capables d’occuper la même fonction dans un réseau de pollinisateurs constituent un groupe fonctionnel.
Exemple d’un papillon Heliconius hecale se nourrissant de nectar sur une fleur de Psychotria (Panama – B. Gilles)
Ce sont ces groupes qui expliquent la résilience des réseaux de pollinisation : si une espèce disparaît, le service est toujours assuré par les autres espèces de son groupe fonctionnel. Ce n’est que lorsque un groupe fonctionnel dans son entier disparaît que des risques d’effondrement des communautés de plantes deviennent réels. Le concept est donc très pertinent pour comprendre la réaction des réseaux de pollinisation face aux perturbations qu’ils subissent.
Considérés au niveau des espèces, les syndromes de pollinisation font débat, nous l’avons vu. Considérés au niveau des groupes fonctionnels de pollinisateurs, c’est à dire de butineurs partageant les mêmes grandes caractéristiques comportementales et morphologiques, ils apparaissent comme un compromis entre généralisme et spécialisation.
Mouche Eristalis tenax – Syrphidae (Source : V. Albouy)
Les traits floraux particuliers qui caractérisent chaque syndrome tendent vers la spécialisation, pour garantir une pollinisation plus efficace en facilitant le travail du pollinisateur. Mais ils restent suffisamment généralistes pour attirer un ensemble plus ou moins vaste d’espèces pollinisatrices, ce qui garantit une meilleure sécurité de la pollinisation en cas de raréfaction ou de disparition des pollinisateurs les plus efficaces. Des « roues de secours » sont ainsi prêtes à être activées. Nous avons vu à l’œuvre un tel mécanisme dans le cas du lierre grimpant où une guêpe peut remplacer une abeille comme pollinisateur efficace.
Généralisme et spécialisation ne peuvent pas être jugés en eux-mêmes mais doivent être appréciés à la lumière des interactions de l’espèce considérée avec son environnement. Ces interactions varient constamment au fil du temps ou des lieux, rebattant sans cesse les cartes. Par exemple l’absence d’un pollinisateur efficace peut être due à une perturbation du milieu naturel, à l’homme ou bien à la petite taille de la population de la plante devenant moins attractive. La petite taille de la communauté des pollinisateurs modifie aussi fatalement ces interactions, comme dans les îles océaniques où certaines plantes généralistes sur le continent voisin deviennent spécialistes par la force des choses, des pans entiers du cortège normal de leurs pollinisateurs étant absents.
La présence de plantes ou d’animaux supplémentaires provoque au final les mêmes effets. Ces perturbations sont particulièrement importantes lorsqu’elles sont dues à des espèces exotiques envahissantes. Par exemple la plante envahissante va offrir une grande masse de ressource aux pollinisateurs locaux, qui vont se reporter sur elle et négliger les plantes autochtones. Ou bien l’animal envahissant va fortement diminuer la population de certains pollinisateurs, comme le varroa ou le frelon asiatique qui s’attaquent aux abeilles mellifères. Ces paramètres peuvent varier entre les communautés de plantes et au fil du temps dans chaque communauté, mais tous ont un impact sur les réseaux de pollinisation donc sur les populations de plantes pollinisées par les animaux.
Réseau de pollinisation
Les études de terrain sur les réseaux de pollinisation pour mieux comprendre le généralisme et la spécialisation se heurtent à d’innombrables interactions qui rendent l’analyse des résultats très ardue.
Matricaire (Matricaria officinalis) (Source : V. Albouy)
Une équipe franco-canadienne (Fontaine et al., 2006 – lien) a réalisé une étude expérimentale d’un réseau de pollinisation en appliquant le principe de Descartes qui veut que lorsqu’un problème est trop complexe pour être étudié expérimentalement, il faut le découper en plusieurs parties plus simples. Ces chercheurs ont donc reconstitué dans des serres des réseaux de pollinisation simplifiés. Ils ont constitué une communauté de plantes comprenant trois espèces généralistes (pollen et nectar facile d’accès sur des corolles plates en forme d’assiette) et trois espèces spécialisées (pollen et nectar difficile d’accès au fond d’une corolle en tube). Les trois généralistes choisis sont la matricaire officinale (Matricaria officinalis), le bec-de-grue (Erodium cicutarium) et la ravenelle (Raphanus raphanistrum), les trois spécialistes la mimule (Mimulus guttatus), la luzerne cultivée (Medicago sativa) et le lotier corniculé (Lotus corniculatus).
La communauté de pollinisateurs est pareillement constituée de deux groupes de trois espèces. Le groupe fonctionnel des généralistes à langue longue, dépassant 8 mm de long, comprend le bourdon terrestre (Bombus terrestris), le bourdon des jardins (B. hortorum) et le bourdon des pierres (B. lapidarius). Ils peuvent butiner aussi bien les fleurs plates que les fleurs en tube. Le groupe fonctionnel des spécialistes à langue courte, atteignant au plus 5 mm, comprend le syrphe bâton (Episyrhus balteatus), l’éristale gluante (Eristalis tenax) et un autre syrphe, un sphaérophore (Sphaerophoria sp.). Ils ne peuvent butiner que les fleurs plates, nectar et pollen des fleurs profondes en tube leur sont inaccessibles. Les syrphes sont considérés comme des pollinisateurs généralistes dans la nature car il existe une grande masse de fleurs au nectar facile d’accès. Ils ne deviennent spécialistes que dans le cadre hyper-simplifié de cette étude.
Bourdon des pierres (Bombus terrestris) (Source : V. Albouy)Bourdon terreste (Bombus terrestris) (Source : V. Albouy)Bourdon des jardins (Bombus hortorum) (Source : V. Albouy)
Trois sortes de communauté végétale ont été constituées dans des cages cubiques en voile de nylon : uniquement des fleurs généralistes, uniquement des fleurs spécialisées, et les deux groupes rassemblés. Et chacune de ces communautés a reçu trois communautés de pollinisateurs différentes : soit uniquement des généralistes, soit uniquement des spécialistes, et les deux groupes rassemblés. L’étude a duré deux ans, en contrôlant le nombre de visites des insectes sur les plantes, ainsi que le succès reproducteur des plantes en se basant sur leur production de fruits et de graines et sur le nombre de plantules poussant après ces deux ans.
Les communautés composées uniquement de fleurs généralistes, au nectar et au pollen facilement accessible, ont connu le même succès reproducteur quelle que soit la communauté de pollinisateur. Leur réseau de pollinisation est suffisamment varié pour supporter la disparition d’un groupe fonctionnel dans son ensemble.
Les communautés composées uniquement de fleurs spécialisées, au nectar et au pollen cachés au fond d’un tube profond, connaissent un succès reproducteur bien plus élevé quand elles sont butinées par les généralistes à langue longue. Lorsque la communauté des pollinisateurs est réduite au seul groupe fonctionnel des spécialistes à langue courte, ce succès est bien plus faible. Le groupe fonctionnel des butineurs à langue longue apparaît donc dans le cas de ces communautés comme un groupe « clef de voûte » : qu’il disparaisse et la communauté s’effondre.
Une analyse plus fine montre que les fleurs généralistes, quand elles sont butinées uniquement par les bourdons, se reproduisent un peu moins bien quand elles sont mélangées avec des fleurs spécialisées que quand elles sont toutes seules. Dans ce dernier cas, les bourdons n’ont pas le choix et les butinent avec assiduité. Par contre, lorsqu’ils ont le choix, ils préfèrent les fleurs profondes. Dans la compétition entre fleurs plates et fleurs profondes pour attirer les bourdons, ces dernières gagnent haut la main.
En fait, et ce n’est pas une surprise, le succès reproducteur des plantes est maximal lorsque spécialistes et généralistes sont mélangés aussi bien dans la communauté végétale que dans le réseau des pollinisateurs. Dans ce cas, les pollinisateurs se partagent le travail : les syrphes se concentrent sur les fleurs plates, et les bourdons sur les fleurs en tube. Chacun au final, plantes comme insectes, se comporte comme un spécialiste, et ce mini-écosystème atteint alors sa plus grande productivité.
Sources
Albrecht M, Schmid B, Hautier Y & Müller CB (2012) : Diverse pollinator communities enhance plant reproductive success. Procedings of the Royal Society B 279 : 4845–4852 (lien)
Bradshaw Jr HD & Schemske DW (2003) : Allele substitution at a flower colour locus produces a pollinator shift in monkeyflowers. Nature 426 : 176–178 (lien)
Fenster CB, Armbruster WS, Wilson P, Dudash MR & Thomson JD (2004) : Pollination syndromes and floral specialization. Annual Review of Ecology, Evolution, and Systematics 35: 375–403 (lien)
Fontaine C, Dajoz I, Meriguet J & Loreau M (2006) : Functional diversity of plant-pollinator interaction webs enhances the persistence of plant communities. PLoS Biol, 4(1), e1 (lien)
Ollerton J, Alarcón R, Waser NM, Price MV, Watts S, Cranmer L, Hingston A, Peter CI & Rotenberry R (2009) : A global test of the pollination syndrome hypothesis. Annals of Botany 103(9) : 1471–1480 (lien)
Ollerton J, Killick A, Lamborn E, Watts S & Whiston M (2007) : Multiple meanings and modes: on the many ways to be a generalist flower. Taxon 56(3) : 717–728 (lien)
Song Z & Feldman MW (2014) : Adaptive foraging behaviour of individual pollinators and the coexistence of co-flowering plants. Proc. R. Soc. B 281: 20132437 (lien)
Philippe Garcelon sur le terrain photographiant un collembole (Source : Corinne Rolland)
Je m’appelle Philippe Garcelon et réside aujourd’hui en région toulousaine. Je suis né dans un petit village du Cantal et dans les années 1960-70, une bonne part de mes distractions se résumait à des escapades dans la nature. Déjà, je débutais mes premières collections d’insectes.
Malheureusement je délaisserai mon « terrain de jeu » pour des études supérieures à l’issue desquelles j’ai intègré un grand groupe français. A cinquante ans, alors cadre de direction, je décide de changer de vie pour me consacrer à des centres d’intérets plus proches de mes aspirations.
Dans un premier temps je me tourne vers l’astronomie et l’astrophysique, ce qui me permet aujourd’hui d’effectuer des missions régulières sur le coronographe du Pic du Midi. Mais je n’en oublie pas pour autant les insectes ou les arachnides qui sont un excellent prétexte pour m’adonner à la macrophotographie que je découvre.
C’est fortuitement, qu’en 2016, je rencontre mes premiers collemboles alors que j’ignorais jusqu’à leur existence. Je cherche à me documenter à leur sujet et constate rapidement la relative pauvreté de publications abordables les concernant. Rapidement, ces petits arthropodes me fascinent, au point que j’éprouve le besoin de partager cet engouement.
Bien sûr, je ne saurais revendiquer la compétence de l’entomologiste de métier pour proposer autre chose qu’une approche vulgarisatrice à leur sujet. Pour autant, je fonde une certaine légitimité de mon propos à travers un noble moteur qui m’anime, celui de la passion.
Ou vivent les collemboles ?
Si méconnus, ils sont pourtant disséminés un peu partout sur Terre. Ils vivent au sol, sur l’eau en bordure des étangs, en haute montagne, dans les déserts, les grottes et même sous des latitudes polaires. Actuellement on en a répertorié plus de 8000 espèces et il en reste probablement davantage à découvrir. On ne soupçonne pas l’abondance de ces créatures. Dans certains habitats, leur densité peut atteindre plusieurs centaines de millions d’individus par hectare, toutes espèces confondues. Par exemple, lorsqu’on se promène en milieu naturel, sous l’empreinte d’un seul de nos pas, ils sont parfois plusieurs centaines à évoluer entre la surface et quelques centimètres de profondeur. Dans les habitats forestiers où abondent les feuilles mortes en décomposition, il n’est pas forcément exceptionnel (dans certaines zones où domine notamment le chêne) de trouver de 200 000 à 300 000 collemboles par m² !
Quelles sont les premières traces écrites à propos des les collemboles ?
Les collemboles sont absents dans la littérature antique. Il semble qu’Aristote (-384 à -322 av. J.-C.) fut le premier à évoquer des « vers » qui pourraient bien être des collemboles. Je le cite : « La neige produit elle-même des vers en vieillissant. Comme le temps la rend rouge, ces vers sont rouges aussi : ils sont velus. En Médie ils sont blancs et grands. Les vers de la neige sont tous paresseux à se mouvoir ». Ensuite, Il faut patienter plus trois siècles avant de retrouver une mention indirecte au texte d’Aristote, évoqué succinctement par Pline l’ancien (23-79) dans son ouvrage « Histoire naturelle ».
Les premières mentions plausibles de cet arthropode, alors décrit comme un « ver », datent seulement de la fin du XVIIème siècle. Par exemple, dans les éphémérides de « l’Académie des curieux de la nature d’Allemagne », le Docteur Charles Raygerus (médecin hongrois) raconte comment en 1672 « Il tomba dans un pré en Hongrie plus de dix espèces de Vers avec de la neige, lesquels y vécurent, & qu’on y vit remper par troupes pendant plus de quatre jours … ».
Apparition du terme « Podura » dans les premières classification de Linné (Source : P. Garcelon)
Quelques années plus tard, un certain Docteur Jean Chrétien Frommannus observe le 2 décembre 1684 des « puces de neige ». Mais Ce n’est qu’au début du XXème siècle que des chercheurs étudieront sérieusement ces animaux auparavant classés en tant qu’insectes. On voit alors apparaitre dans les premières classifications (vers 1740) de Carl von Linné le terme de « Podura » au sein des « Aptera » (signifiant sans ailes) et décrits de la sorte : « courbés et danseurs avec une queue fourchue»
Ou se situent-ils dans la classification ?
Longtemps considérés comme des insectes, les collemboles forment aujourd’hui une classe à part entière au sein du sous embranchement des hexapodes comme le montre ce tableau.
Place des collemboles dans la classification des animaux ( Source : P. Garcelon)
Au sein des quatre ordres, les espèces actuellement identifiées se répartissent de la manière suivante :
Proportion en espèces des différentes familles de collemboles* ( Source : P. Garcelon)
*Données actualisées au 5-11-2017, car on découvre très régulièrement de nouvelles espèces, en particulier avec les études du code barre ADN, permettant de dissocier des espèces sur des sujets morphologiquement très proches
Comment peut-on les observer ?
Généralement, il faut se mettre à quatre pattes, en sous-bois, sur un tapis de feuilles mortes et scruter le sol. Minuscules (0.25 à 4.5 mm), ils cohabitent avec d’autres animaux plus petits comme les acariens. Si on ne voit rien à la première approche, il faut alors soulever délicatement les couches superficielles de la litière et se remettre à observer. Beaucoup d’entre eux sautent lorsqu’ils se sentent en danger, ce qui nuit à l’observation mais, en contrepartie, facilite leur repérage.
Comment les photographier ?
Photographier d’aussi petits animaux, surtout lorsqu’on souhaite le faire en milieu naturel, nécessite l’utilisation d’un minimum de matériel et surtout une bonne connaissance de leur biotope. Pour ne pas être rebuté, il convient de chercher les plus grands (Orchesella – env. 4.5 mm) photographiables avec un objectif macro classique. Mais, très rapidement, on se confronte à l’écueil dimentionnel et aux limites techniques de notre matériel. En effet, Orchesella, qui est déjà un très petit sujet, est encore dix fois plus grand qu’une bonne part des autres collemboles qui mesurent de l’ordre du millimètre ou moins encore (0,25 mm pour les plus petits). L’acquisition de bagues allonge ou d’un multiplicateur de focale devient incontournable. Mais la meilleure solution est d’utiliser l’objectif CANON 65MM MP-E F/2.8 MACRO, unique en son genre et qui permet un rapport de grossissement x5. La contrepartie inévitable est qu’à des grossissements (x3 ou x5) avec, par exemple, une ouverture f/8, les profondeurs de champ sont réduites respectivement à 0.25 mm et 1.03 mm. Autrement dit, la maîtrise d’un tel objectif n’est pas une mince affaire. Mais tel est le prix à payer pour approcher cet autre univers, celui de la micro-photographie.
Comment situer les collemboles sur l’échelle des temps géologique ?
Ils sont parmi les premiers arthropodes à avoir colonisé la planète il y a 400 Millions d’années. La Terre était une étendue d’eau d’où émergeaient quelques blocs groupés autour d’une masse continentale nommé Gondwana. C’est au cours du Silurien et du Dévonien que les plantes chlorophylliennes et plusieurs grands groupes animaux (principalement les Arthropodes et les Vertébrés) ont acquis des structures anatomiques et des fonctions physiologiques qui leur permettent de vivre hors de l’eau et de conquérir les terres émergées. Les vastes espaces marécageux et les zones de faibles profondeurs se peuplent ainsi de nouvelles espèces d’amphibiens. Sur la Terre, parmi les bactéries bien implantées émergent également les arthropodes, comme les crustacés isopodes (cloportes), les myriapodes (mille pattes), les chélicérates (ancêtres des araignées et scorpions), quelques insectes et surtout les premiers collemboles. Les premiers fossiles connus (ci-dessous) classifiés comme collemboles (Rhyniella praeccursor) ont été découverts en 1926 à Rhynie non loin d’Aberdeen en Ecosse, dans des dépôts de vieux grès rouges du Dévonien moyen (env. -380 Ma), par Hirst & Maulik qui les ont apparentés aux Poduromorphes actuels.
Premiers fossiles de collemboles : Rhyniella praeccursor (-380 Ma) – Ecosse ( Source : P. Garcelon)
Depuis cette période, ils ont survécu à des extinctions massives comme celle du Crétacé (-66 Ma) qui a vu disparaître les grands dinosaures. On en retrouve ainsi dans des ambres fossiles de la Baltique, datant de -40 Ma et le plus surprenant est que ces derniers ressemblent presque traits pour traits à des espèces actuelles. Les observer revient donc à effectuer une sorte de voyage dans le temps.
Morphologie
D’apparences diverses, leurs fonctions physiologiques sont similaires et ils disposent de caractères communs : Ils sont entognathes (pièces buccales rentrées dans la tête) ce qui les différencie des insectes, possèdent des antennes segmentées, 3 paires de pattes rattachées aux 3 segments thoraciques et 6 segments abdominaux. Leur corps peut être couvert de poils ou d’écailles. Leur peau possède des propriétés hydrophobes, comme nos tissus déperlants. Ils peuvent présenter des motifs ou diverses pigmentations.
Il existe quatre ordres de collemboles :
Les Entomobryomorphes vivant en surface, dotés d’un organe sauteur nommé furca et dont le corps allongé possède des téguments plus durs couverts de poils
Les Poduromorphes, qui vivent plutôt sous terre, ne sautent pas, sont parfois aveugles et ont un corps mou et souvent boudiné
Les Symphypléones, sauteurs aux segments thoraciques et abdominaux fusionnés leur donnant un aspect globuleux
LesNeelipleones (ordre rassemblant des collemboles « globuleux » de très petit taille)
Description morphologique des différentes familles de collemboles ( Source : P. Garcelon )
Mode de vie
Leur physiologie nécessite un fort taux d’humidité, ils vivent et se reproduisent d’autant mieux que l’hygrométrie est élevée. Il est donc plus aisé de les voir évoluer à la fin de l’automne en hiver ou au printemps que durant les périodes sèches où ils se réfugient profondément et limitent leurs mouvements. Leur durée de vie à l’état naturel est d’environ un an. Leur alimentation est constituée de végétaux en décomposition et peut également comporter des micro-organismes ou des champignons. Leur reproduction peut être sexuée ou asexuée (parthénogénèse).
Reproduction
Une part d’entre eux se livre à des sortes de parades amoureuses (voir vidéo ici : parades amoureuses ) qui rappellent des chorégraphies codifiées selon les espèces et qui ont toujours pour objet l’efficience de la reproduction. Suivant les conditions climatiques, les œufs mettent d’une à quatre semaines pour éclore. Ils ne connaissent pas d’état larvaire, les petits collemboles naissent déjà formés avec la même morphologie que leurs parents. Leur taille évoluera régulièrement par l’intermédiaire de mues successives.
Quelques spécificités anatomiques
La Furca organe sauteur
Organe permettant aux collemboles de réaliser des sauts : la furca – 1) manubrium 2) dentes 3) mucron ( Source : P. Garcelon)
Contrairement aux insectes, ils ne possèdent pas d’ailes. En revanche leur anatomie comporte des organes qui leurs sont propres, comme la « furca » qui leur sert à effectuer des sauts pour fuir des prédateurs ou pour quitter rapidement le milieu où ils évoluent lorsqu’ils sentent un danger. Bien que pouvant dans certains cas maîtriser la direction dans laquelle il saute, le collembole ne maîtrise généralement pas toutes les composantes de ses sauts et, de ce fait, il ne peut pas s’en servir comme moyen ordinaire de locomotion. Sur l’image ci-dessus, la furca, fixée au quatrième segment abdominal est composée, à sa base, du manubrium (1) qui renferme la partie musculaire et qui se divise en deux branches chacune constituée des dentes (2) terminés chacun par un article nommé mucron (3).
Le collophore
Le collophore et sa paire de vésicules excrétiles (Source : P. Garcelon)
Le collembole est également doté d’un tube ventral nommé collophore, sorte de tuyau plus ou moins allongé selon les groupe qui est constitué d’un tube portant à sa base une paire de vésicules excrétiles à parois minces. Ces derniers se rétractent dans le cylindre par l’intermédiaire de douze muscles dits rétracteurs. Ils sont dotés d’organes sensitifs interprétés comme hydrorécepteurs, osmorécepteurs et récepteur pH (acidité/basicité) ainsi l’animal peut augmenter ou réduire ces valeurs en sortant ou en rétractant ses vésicules. La fonction principale du collophore est donc hydrique, à travers le transport de l’eau et des ions vers l’hémolymphe. Cet organe servirait enfin à des fonctions secondaires, comme par exemple, grâce à une glande adhésive, à maintenir le collembole en place lorsqu’il évolue sur des surfaces lisses. On peut également citer la fonction respiratoire grâce au passage de l’oxygène à travers sa paroi fine.
La cuticule
La cuticule (peau) présente également des caractéristiques étonnantes. L’étude des fonctions physiologiques du collembole nous apprend qu’il respire à travers sa cuticule et qu’un dessèchement de cette dernière lui serait fatal, c’est pourquoi il affectionne les milieux humides. Mais l’excès d’humidité lui serait tout aussi fatal s’il venait à recouvrir sa cuticule. C’est donc essentiellement pour se protéger du risque d’asphyxie que le collembole a développé une cuticule dont la nanostructure possède une organisation complexe qui lui permet de repousser efficacement l’eau. Afin de permettre une bonne respiration cutanée, la cuticule doit en effet rester propre, sèche et exempte de toute souillure qui pourrait réduire sa surface fonctionnelle.
Structure de l’épicuticule de la surface d’un collembole en forme de trame de cellules ouvertes ( Source : P. Garcelon)
Exemple : Les propriétés hydrophobes de la cuticule du collembole sont mises en évidence sur cette photographie d’un Dicyrtomina ornata (On doit ici prendre en compte l’évaporation de l’eau contenue dans le substrat et l’accroissement du taux d’humidité de l’air qui s’en suit. Au contact d’une surface froide cette humidité se condense sous la forme de fines gouttelettes, comme on peut le voir dans la nature sur divers supports : herbes, mousses, toiles d’araignées etc…).
Goutelettes à la surface de la cuticule chez Dicyrtomina ornata (Source : P. Garcelon)
Explication : Dans le cas de ce collembole, la chose n’est pas aussi simple car ce n’est pas en raison de la seule condensation que l’on observe la formation des gouttelettes. En effet, la structure de l’épicuticule du collembole forme une trame de cellules ouvertes (revoir le schéma ci-dessus) dans laquelle l’air humidifié se retrouve piégé. L’absence de flux d’air au sein de cette structure aura pour conséquence d’accroître le taux d’humidité qui, devenant supérieur à celui de l’air ambiant, permet la formation de condensation ; non pas en raison d’une différence de température mais bien à cause du différentiel de taux d’humidité, car à température constante, le point de rosée (formation des gouttelettes) dépend du différentiel de saturation de l’air en eau (pour comprendre ce phénomène, voir : point de rosée).
On pourra donc observer, sous certaines conditions, la formation de ces fines gouttelettes dues à la condensation d’une part et, d’autre part, aux propriétés hydrophobes de la cuticule (évoquées précédemment). Les piliers cuticulaires, de par leur structure et aussi grâce à leur surface riche en lipides, rejettent en effet ces gouttelettes en surface. Les plus fines d’entre elles s’agglomèrent ensuite pour former des sphères liquides de plus en plus grosses, jusqu’à ce que la gravité les entraîne vers le sol.
Existe-t-il des espèces possédant des spécificités ?
Collembole Cyphoderus albinus – espèce vivant complémentarité avec les fourmis (Source : P. Garcelon)
Si je me réfère à mes propres observations, je ne pourrais guère citer que le Cyphoderus albinus qui présente la particularité de vivre en parfaite complémentarité avec certaines fourmis. En effet, en échange de travaux de nettoyage (ces collemboles mangent les champignons susceptibles de contaminer les maternités souterraines des fourmis), les fourmis marquent ce collembole avec les phéromones de leur communauté, de telle sorte que ces dernières les considèrent comme des leurs.
Certains Poduromorphes de l’ordre des Neanurinae, comme le Neanura muscorum (image ci-dessous) bien que faciles à prédater en apparence, de par leur morphologie et leurs faibles capacité défensives semblent totalement délaissés par les acariens, araignées ou myriapodes qui sont leurs prédateurs habituels.
Les chercheurs qui ont effectué des études chimio-taxonomiques ont découvert que ces espèces sécrètent des substances répulsives (ici le Dimethoxybenzène) qui suffisent amplement à décourager les prédateurs. Ces recherches effectuées sur de nombreuses autres espèces ont permis de mettre en évidence des évolutions de stratégies défensives directement liée avec les intensités prédatrices qui pèsent sur elles.
Collembole Neanura muscorum – espèce sécrétant des molécules répulsives (Source : P. Garcelon)
Quel rôle jouent les collemboles ?
I) Décomposeurs des sols
Ils jouent un rôle essentiel dans la décomposition de la matière organique, contribuant à l’équilibre écologique de la microfaune en permettant la dissémination et la régulation des populations de champignons et de bactéries. Indispensables à la circulation des nutriments vitaux pour les plantes, ils jouent en amont un rôle dans le découpage et la fragmentation des végétaux avant que les micro-organismes (bactéries, champignons, protozoaires, algues, nématodes…) n’interviennent.
Différentes étapes de la fragmentation organique d’une feuille (Source : P. Garcelon)
On note différentes phases dans le cycle fragmentation organique à travers l’illustration ci-dessus. A chacune des étapes les collemboles jouent un rôle majeur. On peut en établir une chronologie :
D’abord, les feuilles tombées de l’arbre s’entassent dans un premier temps sur le sol où elles sont d’abord lavées par les pluies. La microfaune entre en action. Les arthropodes (collemboles, acariens) commencent à attaquer l’épiderme des feuilles où les premiers trous ne tarderons pas à apparaître
Les petites larves de diptères agrandissent les échancrures et la microflore (bactéries et champignons) entre en action
Les arthropodes continuent de découper les feuilles, les plus robustes d’entre eux attaquent leurs nervures ramollies par la microflore
La fragmentation des débris se poursuit et une partie d’entre eux se mêle aux déjections issues des étapes précédentes
La dissémination des résidus organiques et l’humidité automnale ambiante stimulent la microflore et accélère la dispersion des éléments minéraux
On peut déplorer qu’au terme de ce cycle, les composés chimiques non assimilables par ces organismes vivants, s’ils ne les tuent pas, hypothèquent dangereusement les cycles de biodégradation à venir. C’est ainsi qu’au fil du temps se forment des concentrations croissantes de substances nocives dans les sols. Toute action qui vise à la préservation de la biodiversité ne trouve pas sa seule justification dans des considérations idéologiques, mais plus simplement dans une manifestation de l’instinct de survie. Ainsi, l’homme qui répand des insecticides ou autres poisons ciblant animaux et végétaux parasites (substances pudiquement nommées phytosanitaires) n’en tire qu’un bénéfice à très court terme. En revanche, il peut être assuré que ses actes auront des conséquences dont il sera un jour victime.
II) Marqueur écologique
Dans les études de qualité des sols, les collemboles sont de précieux auxiliaires. La « star » des laboratoires, le Folsomia candida est devenu une espèce standard faisant l’objet d’une norme ISO. Utilisé dans de nombreuses expériences ce dernier a conduit à l’établissement de la norme ISO11267 qui détermine la qualité d’un sol en fonction de l’inhibition à la reproduction de cette espèce. Le comportement des collemboles lors des tests de résistance auxquels on les soumet, permet aux chercheurs d’établir des statistiques sur les nuisances causées aux sols par les agressions chimiques dues aux activités de l’homme (pollution aux métaux, pesticides ou déchets agricoles et domestiques).
Taille des populations de Folsomia candida au contact du « carbaryl », un insecticide et un régulateur de croissance (Source : Filipe & Cardoso, 2002)
L’exemple ci-contre montre le comportement des Folsomia candida au contact du « carbaryl », un insecticide et un régulateur de croissance utilisé dans l’agriculture pour freiner la chute des fruits.
Ici, deux types de population sont testés : Les juvéniles et les adultes, sur une plage de concentration en carbaryl allant de zéro à 7 mg/kg de substrat durant une période de 28 jours. Comme le montre le graphique, les résultats sont édifiants. Avec une dose de 6 mg, les populations sont réduites de moitié alors qu’avec 7 mg, ces dernières sont totalement décimées. On peut noter que durant cette exposition les collemboles ont continué de se reproduire ; entre 4 et 7 mg de concentration des différences significatives ont été enregistrées avec une population test (non exposée). En revanche 7 mg/kg représentent un seuil au-delà duquel tout meurt. EC50 (concentration efficace) se situe à 5.1 mg/kg et LC50 (concentration létale) situe à 5.4mg. On comprend dès lors d’où proviennent les préconisations d’utilisation données par les fabricants de tels produits…
Cet exemple souligne les effets néfastes de toutes ces molécules chimiques que l’agriculture utilise. On peut supposer que les impacts sur l’homme auront, à terme, de graves répercussions sur sa santé. Certes les doses de ces substances qui entrent dans nos organismes sont peut-être infinitésimales relativement à notre masse corporelle, mais de même qu’elles s’accumulent dans nos sols, notre organisme les stocke jusqu’à ce qu’un jour, les concentrations atteignent des seuils fatals, à l’image de ceux qui tuent chaque jour des milliers de petits Folsomia candida dans les laboratoires de recherche du monde entier… Et que faisons-nous pendant ce temps-là ?
Une conclusion ?
Il me semble que nous devrions apprendre à rester humbles devant ces « insignifiantes créatures ». En effet, les collemboles nous indiquent une voie dont nous nous écartons trop souvent par nos pratiques ; celle de la préservation des ressources naturelles, sans lesquelles l’extinction massive qui a déjà commencé au XXème siècle pourrait rapidement exposer notre espèce au premier rang, devant notre incapacité à penser et à agir au-delà de la durée d’une génération.
Un grand merci à « Passion Entomologie » pour m’avoir offert l’occasion d’évoquer les collemboles. Si vous souhaitez en savoir davantage à leur sujet je vous propose de consulter le site que je leur dédie https://collemboles.fr
Source
Filipe D. & Cardoso N. (2002) : Combined effects of carbaryl and abiotic factors to Folsomia candida. Universidad de Aveiro – Departemento de Biologica (lien)
http://www.collembola.org/ : Site de référence tenu par Frans Janssens, membre de la Société Royale d’Entomologie de Anvers. Associé de laboratoire du Département de biologie, groupe de l’écologie évolutive de l’université d’Anvers en Belgique
Seb et Annie, des Criquets Migrateurs, réalisent actuellement un tour du monde pour découvrir la diversité des pratiques culinaires liées l’entomoculture et l’entomophagie. En janvier dernier, Passion-Entomologie les avaient interviewés (lien), les deux voyageurs nous propose ici un nouvel article afin de vous tenir informé de ce qu’ils on pu découvrir jusqu’à présent.
Pour commencer, nous avons sorti toutes les vidéos et tous les articles issus de notre voyage au Japon dont nous vous parlions dans l’article précédent. Nous y revenons sur le déclin et la lutte pour la survie culturelle de la consommation de guêpes, de frelons, de criquets et d’autres bestioles au pays du Soleil Levant !
Au cas où vous en doutiez, tout était parfaitement délicieux !
Cambodge
Nous avons également entamé le gros de notre voyage, un tour du monde de cinq mois qui nous a emmenés au Cambodge et en Thaïlande et qui nous fera bientôt partir en Australie et au Mexique !
Au Cambodge, nous avons pu rencontrer les chasseurs, les producteurs, les cuisiniers et les consommateurs d’insectes – et de mygales, elles ne sont pas des insectes mais nous les comptons quand même !
Il y a au Cambodge une diversité d’insectes consommés à laquelle le Japon ne nous avait pas préparés, la culture entomophage est toujours vivante car le pays est toujours très rural et que les communautés rurales ne s’inquiètent pas « d’occidentaliser » leur alimentation. Nous avons ainsi pu goûter plusieurs variétés de grillons, des taupes-grillons, des nymphes de bombyx du mûrier (c’est-à-dire des vers à soie transformés dont on a déjà dévidé le cocon), des punaises d’eau géantes, et des coléoptères.
Mais les insectes ne sont pas une catégorie à part dans l’alimentation cambodgienne : ils sont en quelque sorte inclus dans une plus vaste catégorie de « non-viandes » dans laquelle on retrouve également les mygales, les scorpions et les grenouilles.
La recette reine pour les insectes et assimilés au Cambodge, c’est la friture, qui donne un peu le même goût à tout. À part ça, tout était intéressant à déguster.
Thaïlande
Nous nous trouvons actuellement en Thaïlande, où la situation est un peu différente bien que ce soit un pays voisin du Cambodge. La Thaïlande est en plein développement et transforme sa production d’insectes : beaucoup de ceux qui sont servis sur les étals sont importés du Cambodge ou d’autres pays environnants. Nous sommes au milieu d’un parcours de visites d’élevages très intéressant qui montre une transition en cours depuis l’insecte chassé dans la nature jusqu’à un insecte semi-domestiqué pour l’alimentation humaine. Nous avons aussi rencontré une des quelques équipes de recherches qui travaillent sur l’entomophagie à travers la Thaïlande ; celle-ci est à la recherche de molécules d’intérêt pour la pharmacologie, l’alimentation ou la cosmétique afin de valoriser les insectes.
Sur les étals de marchés, comme au Cambodge, on trouve des nymphes de bombyx du mûrier, plusieurs espèces de grillons, des punaises d’eau géantes et des coléoptères. Mais on a aussi vu des insectes inédits : vers à soie avant leur transformation en nymphes, criquets géants, œufs de fourmis tisserandes, reines de fourmis tisserandes, vers de bambou, cigales et scarabées rhinocéros – pour le moment. Encore une fois, la friture domine les recettes et nous a sans doute masqué beaucoup de saveurs !
Nous avons encore pas mal de choses à faire ici, puis nous quitterons le continent pour nous rendre en Australie, où nous nous rendrons entre autres dans une communauté aborigène pour en apprendre plus sur la consommation traditionnelle d’insectes du bush. En attendant, vous pouvez d’ores et déjà retrouver les recettes thaïlandaises entomocoles dans cette vidéo culinaire, et toutes les fiches recettes associées :
Toujours pour vous mettre à jour par rapport au dernier article, plusieurs choses ont changé dans notre organisation.
Nous ne pourrons malheureusement pas sortir les épisodes documentaires en même temps que nous voyageons, ils sortiront donc tous en 2019 (si tout va bien, dès janvier ou février !) ; en effet, leur montage représente un travail important qu’il s’est révélé impossible de cumuler avec la découverte, l’écriture et le tournage de tout le reste… Mais promis, on vous abreuvera régulièrement d’articles, de photos et de vidéos culinaires !
Par contre, une bonne nouvelle est que nous avons réuni suffisamment de fonds et de moyens pour que le projet se fasse jusqu’au bout, à ce stade c’est une certitude.
Nous avons changé deux pays de la liste que nous annoncions dans le précédent article : nous ne nous rendons plus au Kenya et au Botswana, mais au Cameroun et au Zimbabwe, qui ont des traditions entomocoles plus ancrées tout en ayant des projets d’élevages d’insectes d’envergure !
Néanmoins, si la communauté des lecteurs et lectrices de Passion-Entomologie veut nous aider, nous recherchons toujours :
Des contacts au Cameroun
Des contacts au Zimbabwe
Pour les suivre en temps réels, les réseaux sociaux :
 Directeur de Recherche émérite au CNRS, Yves Carton a travaillé comme généticien au Laboratoire Evolution, Génomes, Comportement, Ecologie du CNRS et de l’Université Paris-Saclay (EGCE). Depuis plusieurs années, il se consacre à l’histoire des sciences, en particulier dans le domaine de l’entomologie et du darwinisme.
Directeur de Recherche émérite au CNRS, Yves Carton a travaillé comme généticien au Laboratoire Evolution, Génomes, Comportement, Ecologie du CNRS et de l’Université Paris-Saclay (EGCE). Depuis plusieurs années, il se consacre à l’histoire des sciences, en particulier dans le domaine de l’entomologie et du darwinisme.








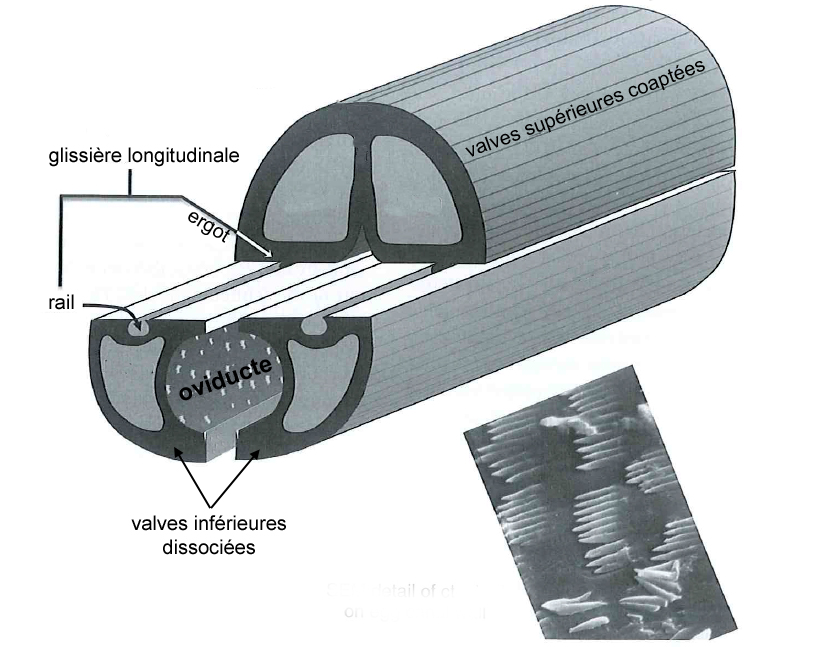





 )
) (N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae – 750 pages – 5 septembre 2013)
(N. Sauvion ; P.A. Calatayud ; D. Thiery & F. Marion-Poll – Editions Quae – 750 pages – 5 septembre 2013) (Bernard Pintureau – Editions : Ellipses Marketing – 188 pages – 25 avril 2009)
(Bernard Pintureau – Editions : Ellipses Marketing – 188 pages – 25 avril 2009) (Vincent Albouy – Editions : Quae Editions – 102 pages – 10 mars 2012)
(Vincent Albouy – Editions : Quae Editions – 102 pages – 10 mars 2012) (Louis M. Schoonhoven, Joop J.A. van Loon & Marcel Dick – Editions : OUP Oxford – 448 pages – 1 décembre 2005)
(Louis M. Schoonhoven, Joop J.A. van Loon & Marcel Dick – Editions : OUP Oxford – 448 pages – 1 décembre 2005)

 (Evrard Wendenbaum – Editions de La Martinière – 173 pages – 17 novembre 2011)
(Evrard Wendenbaum – Editions de La Martinière – 173 pages – 17 novembre 2011) (Evrard Wendenbaum & Pierre Stine – 98mn – 14 décembre 2011)
(Evrard Wendenbaum & Pierre Stine – 98mn – 14 décembre 2011) , IRD Editions/
, IRD Editions/














 (Orin Mcmonigle – Editions Coachwhip Publications – 202 pages – 19 février 2013)
(Orin Mcmonigle – Editions Coachwhip Publications – 202 pages – 19 février 2013)























 (R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)
(R.F. Chapman & Stephen J. Simpson – Edition : Cambridge University Press – 959 pages – 5ème édition : 12 novembre 2012)











 : Sylvian Legeard – Edition « Plume de carotte » – 160 pages – 2012
: Sylvian Legeard – Edition « Plume de carotte » – 160 pages – 2012 : Arnold Van Huis & Jeffery Keith Tomberlin – Edition Wageningen Academic Publishers – 448 pages – 2017
: Arnold Van Huis & Jeffery Keith Tomberlin – Edition Wageningen Academic Publishers – 448 pages – 2017 : Arnold Van Huis, Henk Van Gurp & Marcel Dicke – Columbia University Press – 216 pages – 2014
: Arnold Van Huis, Henk Van Gurp & Marcel Dicke – Columbia University Press – 216 pages – 2014 : Stefan Gates – Ebury Press – 144 pages – 2017
: Stefan Gates – Ebury Press – 144 pages – 2017








 (Damien Laverdunt – Editions Actes Sud Junior – 77 pages – 25 août 2010)
(Damien Laverdunt – Editions Actes Sud Junior – 77 pages – 25 août 2010) (Errol Fuller – Editions Delachaux – 256 pages – 20 février 2014)
(Errol Fuller – Editions Delachaux – 256 pages – 20 février 2014) (Eric Buffetaut – Editions le Cavalier Bleu – 153 pages – 7 juin 2012)
(Eric Buffetaut – Editions le Cavalier Bleu – 153 pages – 7 juin 2012)







 (David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005)
(David Grimaldi & Michael S. Engel – Edition : Cambridge University Press – 772 pages – 16 mai 2005) (P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014)
(P.J. Gullan & P.S. Cranston – Edition : Wiley-Blackwell – 624 pages – 5ème édition : 6 août 2014) (Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009)
(Vincent H. Resh & Ring T. Cardé – Edition : Academic Press – 1024 pages – 2ème édition : 22 juillet 2009) « , Pierre Kerner offre enfin une tribune à des organismes trop souvent méprisés au seul motif qu’ils vivent aux dépens d’autres espèces. Partez à la rencontre d’une multitude de parasites en tout genre, et notamment d’insectes. Ces histoires ne laisseront personne indifférent et raviront les curieux en quête de sensations fortes : entre les vers qui poussent des insectes à se noyer et s’extirpent par leur anus, les tiques qui rendent végétariens ou encore les bébés moules qui vampirisent les poissons, notre monde repousse toujours plus loin les limites de l’extraordinaire et du répugnant !
« , Pierre Kerner offre enfin une tribune à des organismes trop souvent méprisés au seul motif qu’ils vivent aux dépens d’autres espèces. Partez à la rencontre d’une multitude de parasites en tout genre, et notamment d’insectes. Ces histoires ne laisseront personne indifférent et raviront les curieux en quête de sensations fortes : entre les vers qui poussent des insectes à se noyer et s’extirpent par leur anus, les tiques qui rendent végétariens ou encore les bébés moules qui vampirisent les poissons, notre monde repousse toujours plus loin les limites de l’extraordinaire et du répugnant !





















 Auteur de nombreux ouvrages grand public sur la nature en général et les insectes en particulier, il vient de publier «
Auteur de nombreux ouvrages grand public sur la nature en général et les insectes en particulier, il vient de publier «  » aux éditions
» aux éditions 







 (Vincent Albouy & André Fouquet- Editions Delachaux – 96 pages – 26 avril 2018)
(Vincent Albouy & André Fouquet- Editions Delachaux – 96 pages – 26 avril 2018)

 (Vincent Albouy & Claire Felloni – Editions Delachaux – 224 pages – 12 février 2017)
(Vincent Albouy & Claire Felloni – Editions Delachaux – 224 pages – 12 février 2017)
















 (Philippe Lebeaux & Jérôme Cortet – Biotope Editions – 252 pages – 17 novembre 2015)
(Philippe Lebeaux & Jérôme Cortet – Biotope Editions – 252 pages – 17 novembre 2015)